The avian respiratory system delivers oxygen from the air to
the tissues and also removes carbon dioxide. In addition, the
respiratory
system plays an important role in thermoregulation (maintaining normal
body temperature). The avian respiratory system is different from that
of other vertebrates, with birds having relatively small lungs plus
nine
air sacs that play an important role in respiration (but are not
directly
involved in the exchange of gases).
(A). Dorsal view of the trachea (circled) and
the lung of the Ostrich
(Struthio camelus). The lungs are deeply entrenched into the
ribs
on the dorsolateral aspects (arrowhead). Filled circle is on the right
primary bronchus. Note that the right primary bronchus is relatively longer,
rather horizontal and relatively narrower than the left primary bronchus.
Scale bar, 1 cm. (B) Close up of the dorsal aspect of the lung showing
the deep costal sulci (s). Trachea indicated by an open circle; filled circle = right
primary bronchus. Scale bar, 2 cm (Maina and Nathaniel 2001).
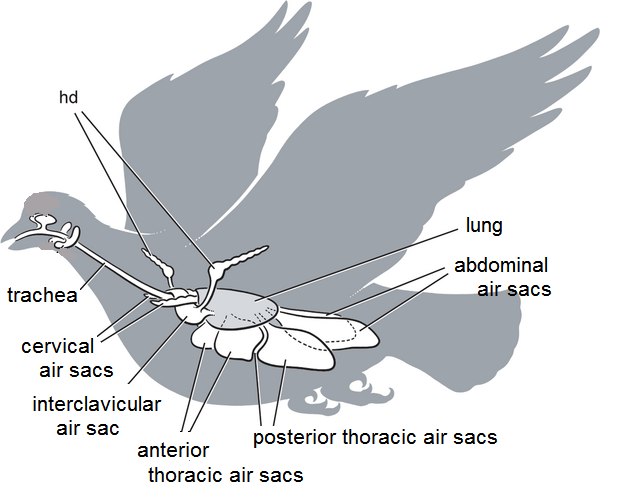
Avian respiratory system
(hd = humeral diverticulum of the clavicular air sac; adapted from
Sereno et al. 2008)
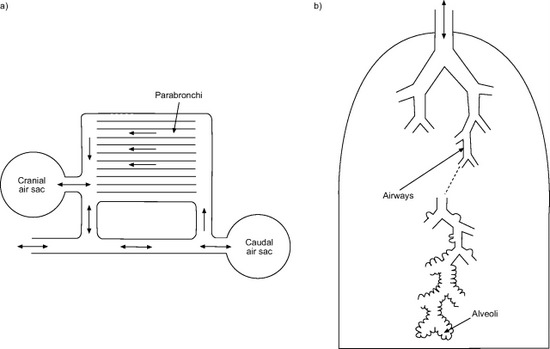
The air sacs permit a unidirectional flow of air through the lungs.
Unidirectional flow means that air moving through bird lungs is largely
'fresh' air & has a higher oxygen content. In contrast, air flow is
'bidirectional' in mammals, moving back and forth into and out of
the
lungs. As a result, air coming into a mammal's lungs is mixed
with
'old' air (air that has been in the lungs for a while) & this
'mixed
air' has less oxygen. So, in bird lungs, more oxygen is available to
diffuse
into the blood (avian
respiratory system).
Pulmonary air-sac system of a Common Teal (Anas crecca). a. Latex injection (blue) highlighting the location of air sacs.
b, Main components of the avian flow-through system. Abd, abdominal aire sac; Cdth, caudal thoracic air sac; Cl, clavicular
air sac; Crth, cranial thoracic air sac; Cv, cervical air sac; Fu, furcula; Hu, humerus; Lu, lung; Lvd, lateral vertebral diverticula;
Pv, pelvis; and Tr, trachea (From: O'Connor and Claessens 2005).
The alveolar lungs of mammals (Rhesus monkey; A) and parabronchial lungs of birds (pigeon; B) are subdivided into large
numbers of extremely small alveoli (A, inset) or air capillaries (radiating from the parabronchi; B, inset). The mammalian respiratory
system is partitioned homogeneously, so the functions of ventilation and gas exchange are shared by alveoli and much of the lung volume.
The avian respiratory system is partitioned heterogeneously, so the functions of ventilation and gas exchange are separate in the air sacs
(shaded in gray) and the parabronchial lung, respectively. Air sacs act as bellows to ventilate the tube-like parabronchi (Powell and Hopkins 2004).
Comparison of the avian 'unidirectional' respiratory system (a) where gases are exchanged between the lungs and the blood in the parabronchi, and the bidirectional respiratory system of mammals (b) where gas exchange occurs in small dead-end sacs called alveoli (From: West et al. 2007).
Avian lungs are supported by a system of air sacs distributed through a birds body that function to generate unidirectional flow of air through the lungs that are subdivided into parabronchi and air capillaries (on the right - red arrows=inspiration; blue arrow=expiration). Magnified inset (below) shows the avian lung subdivided into air capillaries (AC) through which pulmonary capillaries form an enmeshed network with cross-current gas exchange (From: Hawkes 2025).
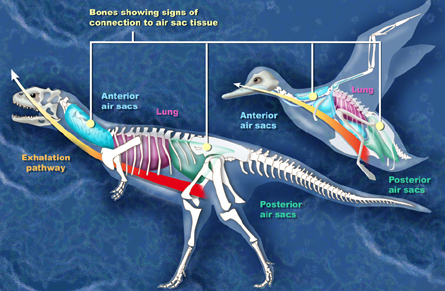
Bird-like
respiratory systems in dinosaurs -- A recent analysis showing
the presence of a very bird-like
pulmonary, or lung, system in predatory dinosaurs provides more
evidence of an evolutionary link between dinosaurs and birds. First
proposed in the late 19th century, theories about the
animals' relatedness enjoyed brief support but soon fell out of favor.
Evidence gathered over the past 30 years has breathed new life
into the hypothesis. O'Connor and Claessens (2005) make clear the
unique pulmonary system of birds, which has
fixed lungs and air sacs that penetrate the skeleton, has an older
history than previously realized. It also dispels the theory that
predatory dinosaurs had lungs similar to living reptiles, like
crocodiles.
The avian pulmonary system uses "flow-through
ventilation," relying
on a set of nine flexible air sacs that act like bellows to move air
through the almost completely rigid lungs. Air sacs do not take part in
the actual oxygen exchange, but do greatly enhance its efficiency and
allow for the high metabolic rates found in birds. This system also
keeps the volume of air in the lung nearly constant. O'Connor says the presence of an
extensive pulmonary air sac system
with flow-through ventilation of the lung suggests this group of
dinosaurs could have maintained a stable and high metabolism, putting
them much closer to a warm-blooded existence. "More and more
characteristics that once defined birds--feathers,
for example--are now known to have been present in dinosaurs, so, many
avian features may really be dinosaurian," said O'Connor. A portion of
the air sac actually integrates with the skeleton,
forming air pockets in otherwise dense bone. The exact function of this
skeletal modification is not completely understood, but one explanation
theorizes the skeletal air pockets evolved to lighten the bone
structure, allowing dinosaurs to walk upright and birds to fly.
Some hollow bones are providing solid new evidence of how birds evolved from dinosaurs.
Most birds have 9 air sacs:
one interclavicular sac
two cervical sacs
two anterior thoracic sacs
two posterior thoracic sacs
two abdominal sacs
Functionally, these 9 air sacs can be divided into anterior sacs
(interclavicular,
cervicals, & anterior thoracics) & posterior sacs (posterior
thoracics
& abdominals). Air sacs have very thin walls with few blood
vessels.
So, they do not play a direct role in gas exchange. Rather, they act as
a 'bellows' to ventilate the lungs (Powell 2000).
Air sacs and axial pneumatization in an extant avian.
The body of bird in left lateral view, showing the cervical (C),
interclavicular
(I), anterior thoracic (AT), posterior thoracic (PT), and abdominal
(AB)
air sacs. The hatched area shows the volume change during exhalation.
The
cervical and anterior thoracic vertebrae are pneumatized by diverticula
of the cervical air sacs. The posterior thoracic vertebrae and
synsacrum
are pneumatized by the abdominal air sacs in most taxa. Diverticula of
the abdominal air sacs usually invade the vertebral column at several
points.
Diverticula often unite when they come into contact, producing a system
of continuous vertebral airways extending from the third cervical
vertebra
to the end of the synsacrum. Modified from Duncker 1971 (Wedel 2003).
Computerized axial
tomogram of an awake, spontaneously
breathing goose; air is darkest. A large percentage of the bird's body
is filled with the several air sacs. Upper left: At the
level
of the shoulder joints (hh, humeral head) is the intraclavicular air
sac
(ICAS), which extends from the heart cranially to the clavicles (i.e.,
furcula or wishbone). S, sternum; FM, large flight muscles with
enclosed
air sac diverticula, arrowheads; t, trachea. Upper right: At
the
level of the caudal heart (H) is the paired cranial thoracic air sacs
(TAS).
Arrowhead points to the medial wall of the air sac (contrast enhanced
with
aerosolized tantalum powder). The dorsal body cavity is filled with the
lungs, which are tightly attached to the dorsal and lateral body wall.
V, thoracic vertebrae. Lower left: At the level of the knees
(K)
is the paired caudal thoracic air sacs (PTAS) and paired abdominal air
sacs, with the abdominal viscera (AV) filling the ventral body cavity.
The membrane separating the abdominal air sacs from one another
(arrowhead)
and from the caudal thoracic air sacs (arrows) can be seen. Lower
right:
At the level of the caudal pelvis, the abdominal air sacs, which extend
to the bird's tail, can be seen. Arrow, membrane separating abdominal
air
sacs (Brown et al. 1997).
Birds can breathe through the mouth or the nostrils (nares). Air
entering
these openings (during inspiration) passes through the pharynx &
then
into the trachea (or windpipe). The trachea is generally as long as the
neck. However, some birds, such as cranes, have an exceptionally long
(up
to 1.5 m) trachea that is coiled within the hollowed keel of the
breastbone
(shown below). This arrangement may give additional resonance to their
loud
calls (check this short video of calling Sandhill Cranes).
Sandhill Cranes calling in flight
Curl-crested Manucode: Unusual Tracheal Anatomy
The typical bird trachea is 2.7 times longer and 1.29 times wider
than
that of similarly-sized mammals. The net effect is that tracheal
resistance
to air flow is similar to that in mammals, but the tracheal dead space
volume is about 4.5 times larger. Birds compensate for the larger
tracheal
dead space by having a relatively larger tidal volume and a lower
respiratory
frequency, approximately one-third that of mammals. These two factors
lessen
the impact of the larger tracheal dead space volume on ventilation.
Thus,
minute tracheal ventilation is only about 1.5 to 1.9 times that of
mammals
(Ludders 2001).
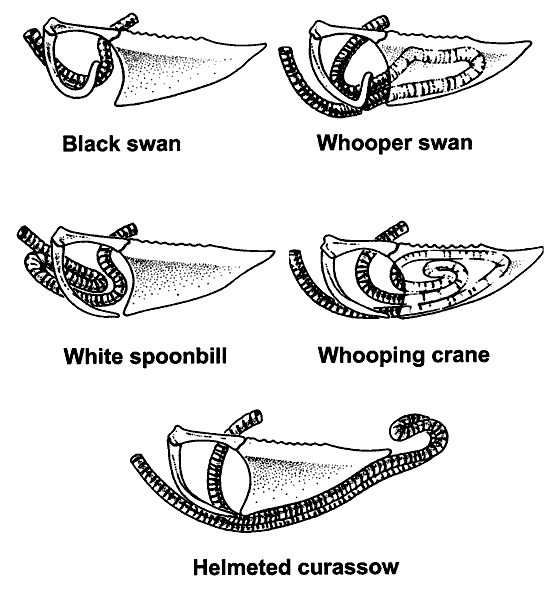
Examples of tracheal loops found in Black Swans (Cygnus
atratus), Whooper
Swans (Cygnus cygnus), White Spoonbills (Platalea leucorodia),
Helmeted Curassow (Crax pauxi),
and Whooping Cranes (Grus americana).
Source: http://www.ivis.org/advances/Anesthesia_Gleed/ludders2/chapter_frm.asp
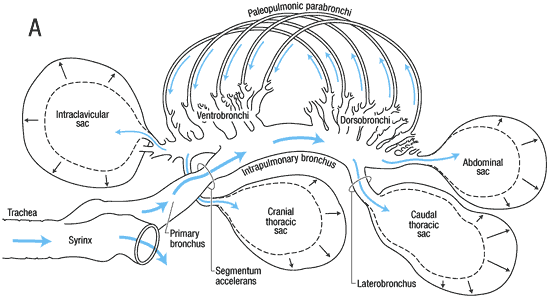
The trachea bifurcates (or splits) into two primary bronchi
at
the syrinx. The syrinx is unique to birds & is their
'voicebox'
(in mammals, sounds are produced in the larynx). The primary
bronchi enter the lungs & are then called mesobronchi.
Branching off from the mesobronchi are smaller tubes called dorsobronchi.
The dorsobronchi, in turn, lead into the still smaller parabronchi. Parabronchi
can
be several millimeters long and 0.5 - 2.0 mm in diameter (depending on
the size of the bird) (Maina 1989) and their walls contain hundreds of
tiny, branching, & anastomosing 'air
capillaries' surrounded by a profuse network of blood
capillaries
(Welty and Baptista 1988).
It is within these 'air capillaries' that the exchange of gases (oxygen
and carbon dioxide) between the lungs and the blood occurs. After passing through the parabronchi, air moves into the ventrobronchi.
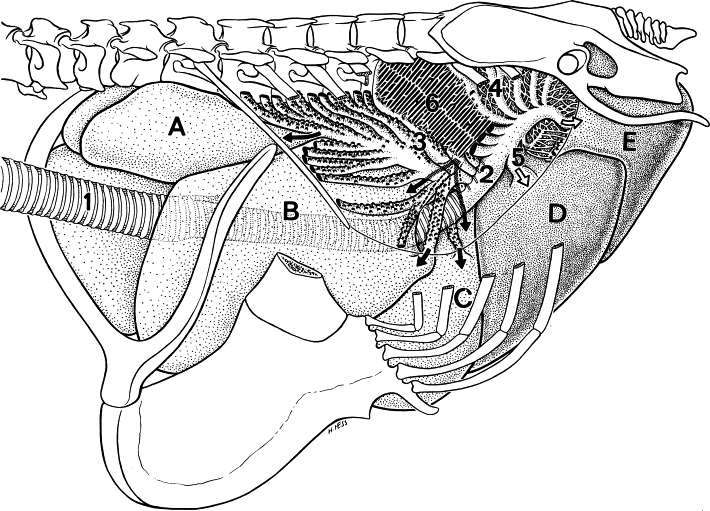
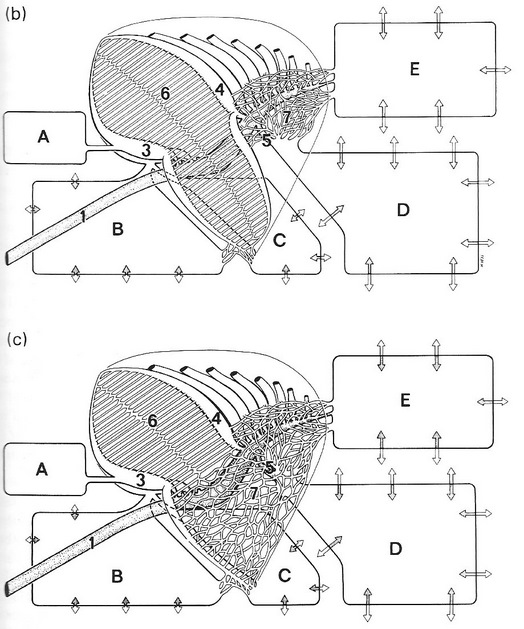
Semi-schematic drawing of the lung-air sac system in situ. The cranial half of the dorsobronchi (4) and the parabronchi (6) has been removed. 1 = trachea, 2 = primary bronchus, 3 = ventrobronchi with the connections into (A) cervical, (B) interclavicular and (C) cranial thoracic air sacs, 5 = laterobronchi and the caudal primary bronchus open into the (D) posterior thoracic and (E) abdominal air sacs (From: Duncker 2004).
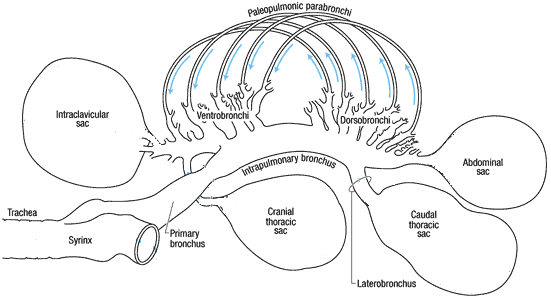
Avian respiratory system showing the bronchi located inside the lungs. Dorsobronchi and ventrobronchi branch off of the primary bronchus; parabronchi extend from the dorsobronchi to the ventrobronchi. Light blue arrows indicate the direction of air flow through the parabronchi. The primary bronchus continues through the lung and opens into the abdominal air sac.
(Source: http://www.ivis.org/advances/Anesthesia_Gleed/ludders2/chapter_frm.asp)
Birds exhibit some variation in lung structure and, specifically, in the arrangement of parabronchi. Most birds have two sets of parabronchi, the paleopulmonic (‘ancient lung’) and neopulmonic (‘new lung’) parabronchi. However, the neopulmonic region is absent in some birds (e.g., penguins) and poorly developed in others (e.g., storks [Ciconiidae] and ducks [Anatidae]). In songbirds (Passeriformes), pigeons (Columbiformes), and gallinaceous birds (Galliformes), the neopulmonic region of the lung is well-developed (Maina 2008). In these latter groups, the neopulmonic parabronchi contain about 15 to 20% of the gas exchange surface of the lungs (Fedde 1998). Whereas airflow through the paleopulmonic parabronchi is unidirectional, airflow through the neopulmonic parabronchi is bidirectional. Parabronchi can be several millimeters long and 0.5 - 2.0 mm in diameter (depending on the size of the bird) (Maina 1989) and their walls contain hundreds of tiny, branching, and anastomosing air capillaries surrounded by a profuse network of blood capillaries.
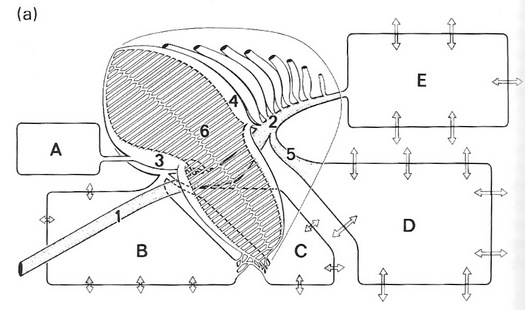
Differences among different birds in the development of the neopulmonic region of the lung. (a) Penguin lungs are entirely paleopulmonic.
(b) Some birds, such as ducks, have a relatively small neopulmonic region. (c) Songbirds have a well-developed neopulmonic region.
1, trachea, 2, primary bronchus, 3, ventrobronchus, 4, dorsobronchus, 5, lateral bronchus, 6, paleopulmonic parabronchi,
7, neopulmonic parabronchi; A, cervical air sac, B, interclavicular air sac, C, cranial thoracic air sac, D, caudal thoracic air sac,
E, abdominal air sac. The white arrows indicate changes in volume of the air sacs during the respiratory cycle (From: McLelland 1989).
So, how does air flow through the avian lungs & air sacs
during respiration?
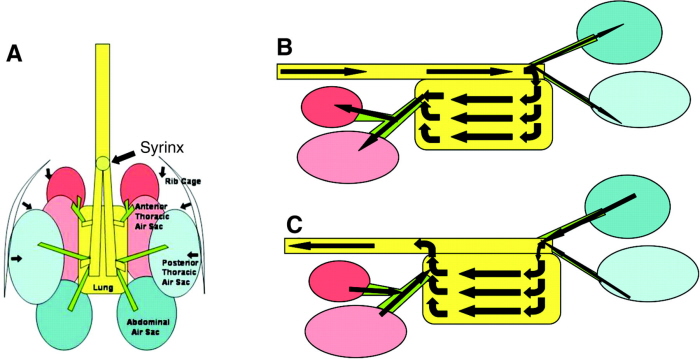
Air flow through the avian respiratory system during inspiration (a) and expiration (b).
1 - interclavicular air sac, 2 - cranial thoracic air sac, 3 - caudal thoracic air sac, 4 - abdominal air sac
(From: Reese et al. 2006).
A schematic of the avian respiratory system, illustrating the major air sacs and their connections to the lung. (A) The lateral and dorsal direction of motion of the rib cage during exhalation is indicated by arrows. (B) The direction of airflow during inspiration. (C) The direction of flow during expiration (From: Plummer and Goller 2008).
Respiratory airflow in avian lungs.Filled and open arrows denote direction of air flow during inspiration (filled arrows) and expiration (open arrows), respectively. Relative thickness of the arrows indicates the proportion of air streaming through the different areas of the respiratory system during the respiratory cycle. Dotted arrows indicate the volume changes of air sacs. In bird lungs (A), most air directly enters the caudal air sacs during inspiration (thick black arrow), whereas a lesser part flows through the parabronchi/air capillaries into cranial air sacs (thin black arrows). During expiration the major part of inspired air streams from the reservoirs (caudal air sacs, thick open arrows) through the parabronchi/air capillaries into major distal airways, where it mixes with the deoxygenated respiratory gas stored in cranial air sacs during the inspiratory phase. Consequently, respiratory gas flow through the parabronchi, atria, and the gas-exchanging air capillaries is unidirectional and continuous during both inspiration and expiration. This principle is achieved by cranio-caudal pressure gradients in the respiratory system changing between inspiration and expiration and the consecutive opening and closing of valve systems between mesobronchi/air sacs and the parabronchi (not indicated in the figure). Hence, airflow is constant and high in the parabronchi, atria, and the gas-exchanging air capillaries (From: Bernhard et al. 2004).
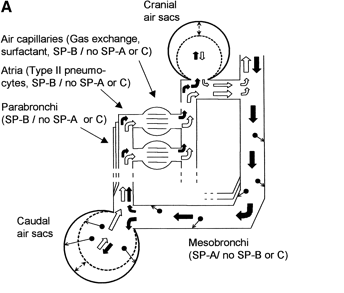
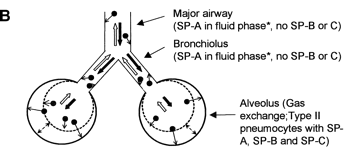
Surfactant SP-B (in the figure above) is mixture of phospholipidsand specific proteins that functions to maintain airflow through the 'tubes' of the avian respiratory system. Surfactant SP-A has only been detected in the mesobronchi of birds. SP-A plays animportant role in innate host defense and regulation of inflammatoryprocesses and may be important in the mesobronchi because air flow is slower and small particles could tend to accumulate there (see figure below). Surfactant SP-C is not found in the avian respiratory system (or, if so, in very small quantities), but is found in the alveoli of mammals along with SP-A and SP-B. Because the mammalian respiratory system (below) includes structures that are collapsible (alveoli) and areas with low airflow, all three surfactants are important for reducing surface tension and innate host defense (Bernhard et al. 2004).
Airflow in mammalian lungs is bidirectional during the respiratory cycle, with highly reduced
airflow
in peripheral structures, i.e., bronchioles and, particularly, the
gas-exchanging alveoli. Consequently, small particles (< 1 µm)
that enter the alveoli may sediment, making a system of first line of defense necessary, comprising alveolar macrophages
(white blood cells), SP-A, and (phospholipid) regulators of inflammatory processes (From: Bernhard et al. 2004).
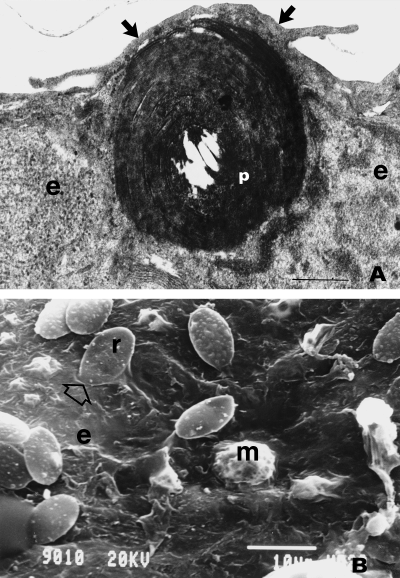
A: A high-power view of a foreign particle (p) being engulfed by an epithelial cell (e) in an avian lung.
Arrows, elongated microvilli. B: Surface of an atrium of the lung of the domestic fowl showing red blood
cells with one
of them (r) being engulfed by the underlying epithelial cell (arrow): e, epithelial surface; m, a free
(surface) macrophage. Scale bars: A = 0.5 µm; B = 10 µm (From: Nganpiep and Maina 2002).
Air flow is driven by changes in pressure within the respiratory
system:
During inspiration:
the sternum moves forward and downward while the vertebral ribs move
cranially to expand the sternal ribs and the thoracoabdominal cavity
(see diagrams below). This expands the posterior and anterior air sacs and lowers the pressure, causing air to move into those
air sacs.
Air from the trachea and bronchi moves into the posterior air sacs
and,
simultaneously,
air from the lungs moves into the anterior air sacs.
Changes in the position of the thoracic skeleton during
breathing in a bird. The solid lines represent
thoracic position at the end of expiration while the
dotted lines show the thoracic position
at the end of inspiration (Source:
http://www.ivis.org/advances/Anesthesia_Gleed/ludders2/chapter_frm.asp).
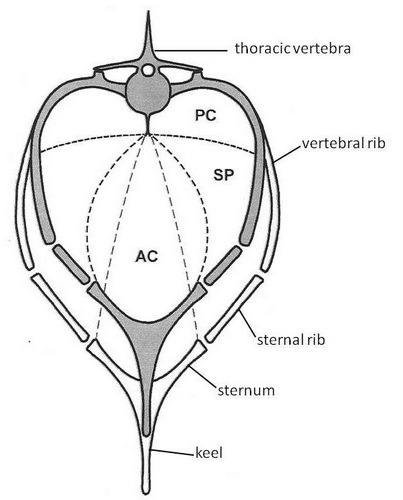
Drawing of a bird coelom in transverse section during expiration (gray bones) and inspiration (white bones). Dashed lines illustrate the
horizontal septum that separates the pleural cavity (PC) where the lungs are located from the subpulmonary cavity (SP) where most
of the air sacs are located (except the abdominals that are in the peritoneal cavity), and the oblique septum that separates the air sacs from
the abdominal cavity (AC) and digestive viscera. Both septa insert on the ventral keel of vertebrae. The volume of the pleural cavity changes
very
little with respiratory rib movements, but the volume of the subpulmonary cavity (and the air sacs) is greatly increased when the oblique
septum is stretched during inspiration (Adapted from: Klein and Owerkowicz 2006). The increase in volume lowers air pressure and draws air
into
the air sacs.
Schematic representation of the lungs
and air sacs of a bird and the pathway of
gas flow through
the pulmonary system during inspiration and expiration. For purposes of
clarity, the neopulmonic lung
is not shown. The
intrapulmonary bronchus is also known as the mesobronchus. A -
Inspiration.
B - Expiration
Source: http://www.ivis.org/advances/Anesthesia_Gleed/ludders2/chapter_frm.asp
During expiration:
the sternum moves backward and upward & the vertebral ribs move
caudally
to retract the sternal ribs and reduce the volume of the
thoracoabdominal
cavity. The reduces the volume of the anterior & posterior air sacs, causing air to move out of those sacs.
Air from the posterior sacs moves into the lungs &, simultaneously,
air from the anterior sacs moves into the trachea & out of the body.
So, air always moves unidirectionally through the lungs and, as a result, is higher in oxygen content than, for example, air in the alveoli of humans and other mammals.
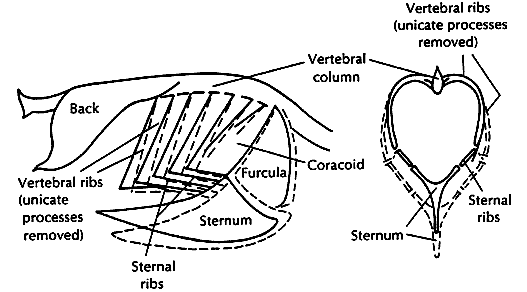
Role of uncinate processes and associated muscles in avian respiration -- Codd et al. (2005) examined the activity of three muscles associated with the uncinate processes, (1) external intercostal, (2) appendicocostalis and (3) external oblique (labeled in drawing to the left) examined using electrodes during sitting, standing and moderate speed treadmill running in a Giant Canada Goose. The external intercostal muscles demonstrated no respiratory activity, being active only during running, suggesting they play some role in trunk stabilization. The appendicocostalis and external oblique muscles are respiratory muscles, being active during inspiration and expiration, respectively. The activity of the appendicocostalis muscle increased when sternal movements were restricted, suggesting that activity of these muscles may be particularly important during prolonged sitting such as during egg incubation. Codd et al. (2005) suggested that the uncinate processes in birds facilitate movements of the ribs and sternum during breathing and therefore are integral to the breathing mechanics of birds.
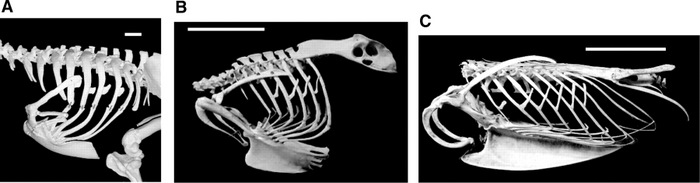
Variation in length of uncinate processes -- Birds with different forms of locomotion exhibit morphological differences in their rib cages: (A) terrestrial (walking) species, Cassowary (Casuaris casuaris); (B) a typical flying bird, Eagle Owl (Bubo bubo); and (C) an aquatic, diving species, Razorbill (Alca torda). Uncinate processes are shorter in walking species, of intermediate length in typical birds, and relatively long in diving species (scale bar, 5 cm). Muscles attached to uncinate processes (appendicocostales muscles) help rotate the ribs forwards, pushing the sternum down and inflating the air sacs during inspiration. Another muscle (external oblique) attached to uncinate processes pulls the ribs backward, moving the sternum upward during expiration. The longer uncinate processes of diving birds are probably related to the greater length of the sternum and the lower angle of the ribs to the backbone and sternum. The insertion of the appendicocostales muscles near the end of the uncinate processes may provide a mechanical advantage for moving the elongated ribs during breathing (Tickle et al. 2007).
Ultra-Low Oxygen Could Have Spurred Bird
Breathing
System -- Recent evidence suggests that oxygen levels were
suppressed
worldwide 175 - 275 million years ago, low enough to make breathing the
air at sea level feel like respiration at high altitude.
Peter
Ward, a University of Washington paleontologist, theorizes that low
oxygen and repeated short but substantial
temperature increases because of greenhouse warming sparked two major
mass-extinction
events. In addition, he believes the conditions spurred the development
of an unusual breathing system in Saurischian dinosaurs. Rather than
having
a diaphragm to force air in and out of lungs, the Saurischians had
lungs
attached to a series of thin-walled air sacs that appear to have
functioned
something like bellows to move air through the body. This breathing
system,
still found in today's birds, made the Saurischian dinosaurs better
equipped
than mammals to survive the harsh conditions in which oxygen content of
air at the Earth's surface was only about half of today's 21%. "The
literature
always said that the reason birds had sacs was so they could breathe
when
they fly. But I don't know of any brontosaurus that could fly," Ward
said.
"However, when we considered that birds fly at altitudes where oxygen
is
significantly lower, we finally put it all together with the fact that
the oxygen level at the surface was only 10 - 11% at the time the
dinosaurs
evolved. That's the same as trying to breathe at 14,000 feet. If you've
ever been at 14,000 feet, you know it's not easy to breathe," he said.
In the avian lung, oxygen diffuses (by simple diffusion) from the air
capillaries into the blood & carbon dioxide from the blood into
the
air capillaries (shown in this figure and in figures below ). This exchange is very efficient in birds for a number of reasons. First, the complex arrangement of blood and air capillaries in the avian lung creates a substantial surface area through which gases can diffuse. The surface area available for exchange (SAE) varies with bird size. For example, the ASE is about 0.17 m 2 for House Sparrows (about 30 gms; Passer domesticus), 0.9 m 2 for Rock Pigeons (about 350 gms; Columba livia), 3.0 m 2 for a Mallard (about 1150 gms; Anas platyrhynchos), and 8.9 m 2 for a male Graylag Goose (about 3.7 kg; Anser anser) (Maina 2008). However, smaller birds have a greater SAE per unit mass than do larger birds. For example, the SAE is about 90 cm 2/gm for Violet-eared Hummingbirds (Colibri coruscans; Dubach 1981), about 26 cm 2/gm for Mallards, and about 5.4 cm 2/gm for Emus (Dromaius novaehollandiae; Maina and King 1989). Among mammals, there is also a negative relationship between SAE and body size, with smaller mammals like shrews having a greater SAE per unit mass than larger mammals. However, for birds and mammals of similar size, the SAE of birds is generally about 15% greater (Maina et al. 1989).
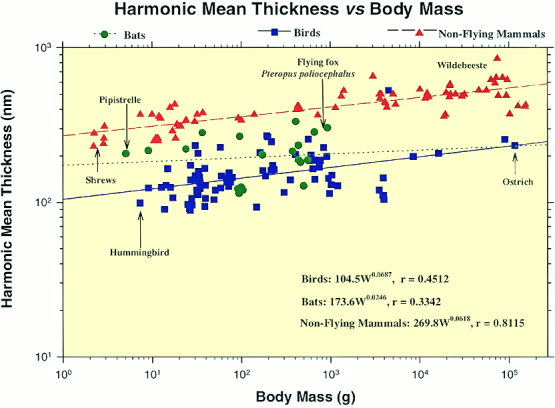
A second reason why gas exchange in avian lungs is so efficient is that the blood-gas barrier through which gases diffuse is extremely thin. This is important because the amount of gas diffusing across this barrier is inversely proportional to its thickness. Among terrestrial vertebrates, the blood-gas barrier is thinnest in birds. Natural selection has favored thinner blood-gas barriers in birds and mammals because endotherms use oxygen at higher rates than ectotherms like amphibians and reptiles. Among birds, the thickness of the blood-gas barrier varies, with smaller birds generally having thinner blood-gas barriers than larger birds. For example, the blood-gas barrier is 0.099 μm thick in Violet-eared Hummingbirds and 0.56 μm thick in Ostriches (West 2009).
Comparison of the mean thickness of the blood-gas barrier of 34 species of birds, 37 species of mammals,
16 species of reptiles, and 10 species of amphibians revealed that birds had significantly thinner blood-gas
barriers than the other taxa (West 2009).
Also contributing to the efficiency of gas exchange in avian lungs is a process called cross-current exchange. Air passing through air capillaries and blood moving through blood capillaries generally travel at right angles to each other in what is called cross-current flow (Figure below; Makanya and Djonov 2009). As a result, oxygen diffuses from the air capillaries into the blood at many points along the length of the parabronchi, resulting in a greater concentration of oxygen (i.e., higher partial pressures) in the blood leaving the lungs than is possible in the alveolar lungs of mammals (Figures below).
Diagram of parabronchial anatomy,
gas-exchange region
of the bird's lung-air-sac respiratory system. The few hundred to
thousand
parabronchi, one of which is fully shown here, are packed tightly into
a hexagonal array. The central parabronchial lumen, through which gas
flows
unidirectionally during both inspiration and expiration is surrounded by gas-exchange tissue composed of an
intertwined
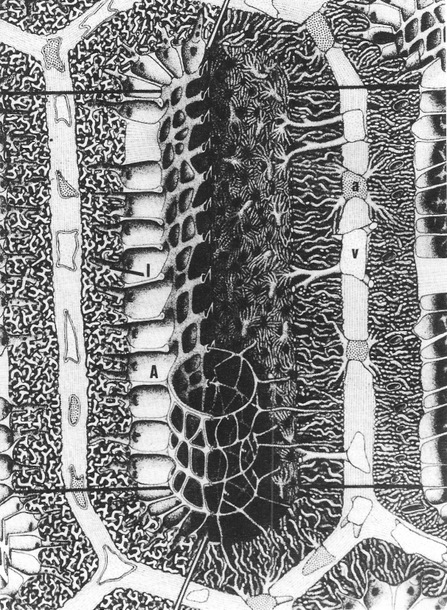
network of blood and air capillaries. On the left side of this diagram, the lumen of the parabronchus leads into multiple chambers called atria (A) that, in turn, lead into smaller chambers called infundibulae (I). Branching from the infundibulae are numerous air capillaries. On the right side of this diagram are the blood vessels. Arteries (a) lead into the capillaries that are closely associated with the air capillaries. It is here (air and blood capillaries) where oxygen and carbon dioxide are exchanged. After flowing through the capillaries, blood then moves into the veins (v) that will take the blood out of the lungs (From: Duncker 1971 as reprinted in Powell 2000).
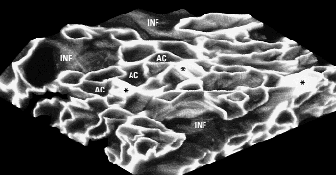
Three-dimensional reconstruction of the gas-exchange region.
AC = air capillaries. Several air capillaries coalesce into an infundibulum (INF) (Brown et al. 1997).
In this cross-section, note the intertwined network
of blood capillaries, labeled with the presence of
erythrocytes (*), and air capillaries (AC) that make up the parabronchi's mantle of gas-exchange tissue (Brown et al. 1997).
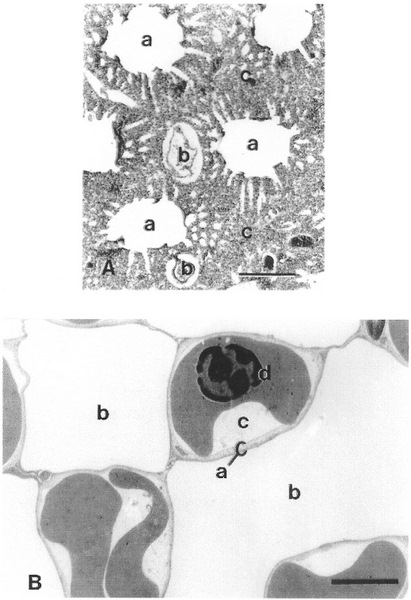
(A) Micrograph of lung tissue from a Brown Honeyeater (Lichmera indistincta) showing (a) parabronchi, (b) blood vessel, and (c) exchange tissue (bar, 200 micrometers). (B) Electron micrograph from the lung of a Welcome Swallow (Hirundo neoxena) showing (a) blood-air barrier, (b) air capillary, (c) blood capillary, and (d) red blood cell in the blood capillary (bar, 2 micrometers). (From: Vitali and Richardson 1998).
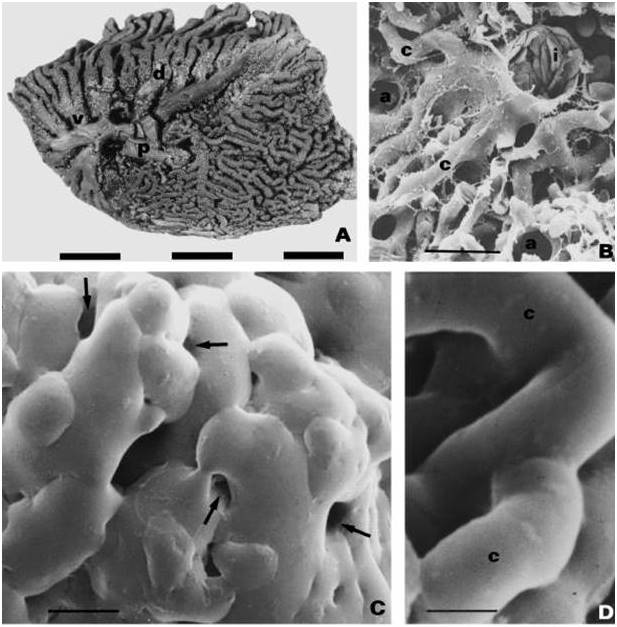
A) Medial view of the lung of a domestic chicken (Gallus gallus domesticus). p, primary bronchus; v, ventrobronchus; d, dorsobronchus; r, parabronchi. Scale bar, 1 cm. (B) An intraparabronchial artery (i) giving rise to blood capillaries (c) in the lung of an Emu (Dromiceus novaehollandiae). a, air capillaries. Scale bar, 15 μm. (C) Air capillaries closely associated with blood capillaries (arrows) in a chicken lung. Scale bar, 10 μm. (D) Blood capillaries (c) closely associated with air capillaries (spaces) in a chicken lung. Scale bar, 12 μm. (From: Maina 2002).
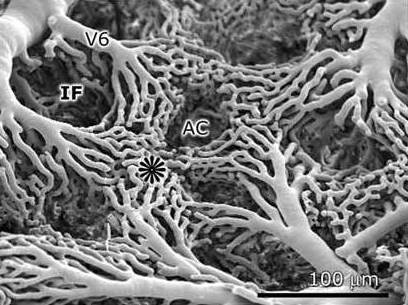
An individual air capillary (AC) surrounded by a dense network of blood
capillaries (asterisk) in a chicken lung. The blood capillaries drain into a
larger vein (V6) adjacent to an infundibulum (IF). Note that the general direction
of
blood
flow through
the blood capillaries is perpendicular to the flow of air through
the air capillaries, i.e., cross-current flow (From: Makanya and Djonov 2009).
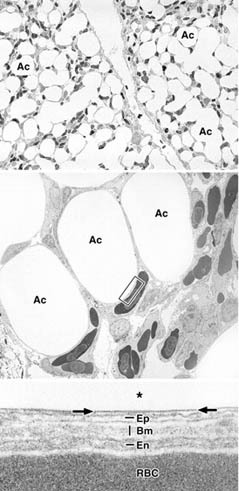
Morphology of a chicken lung. Light microscopy (top image) and electron microscopy (bottom two images) of a chicken lung depicting the respiratory system of birds. In the bird lung, air capillaries (Ac) run along with blood capillaries forming the blood-air barrier that is typically < 0.2 µm in thickness. The barrier (shown in the bottom image) separates the lumen of the Ac (*) from the red blood cells (RBC) in the blood capillaries and consists of a mostly continuous surfactant layer (arrows), thin cytoplasmic processes of epithelial cells (Ep), a common basal membrane (Bm), and the endothelial cells of the blood capillary (En). Surfactant is a mixture of lipids and proteins that acts in the air capillaries of avian lungs both as an "antiglue" (preventing the adhesion of respiratory surfaces that may occur when the lungs collapse, e.g., during diving, swallowing of prey or on expiration) and to prevent liquid influx into the lungs (Daniels et al. 1998). Magnifications: top image - ×270; middle image - ×1,600; bottom image - ×88,000 (Image from Bernhard et al. 2001).
In birds, the thicknessof the blood-gas barrier in the 7.3-g Violet-eared Hummingbird (Colibri coruscans) is 0.099 µm,
whereas that of an immature 40-kg Ostrich (Struthio camelus) is 0.56 µm (Maina and West 2005).
Relationship between the harmonic mean thickness of the blood-gas barrier (the thickness of the barrier that affects the diffusion of oxygen from air capillaries into blood capillaries) against body mass in the lungs of bats, birds, and non-flying mammals. Birds have particularly thinner barriers than bats and non-flying mammals
(Maina 2000).
Light micrographs of a portion of the lung of a chicken (A) and rabbit (B).
Note the small diameter of the air capillaries in the chicken lung vs. that of the rabbit alveoli (same magnification).
(A) In the chicken lung, pulmonary capillaries are supported by 'struts' of epithelium (arrows). (B) In the rabbit lung,
pulmonary capillaries are suspended in the large spaces between alveoli (Watson et al. 2007).
Cross-current exchange:
Top: Air flow (large arrows) and
blood
flow (small arrows) illustrating the cross-current
gas-exchange
mechanism operating
in the avian lung (between the blood capillaries and air capillaries). Note the serial arrangement of
blood
capillaries running from the periphery to the lumen of the parabronchus
and the air capillaries
radially extending from the parabronchial
lumen. The exchange of gases (simple diffusion of O2 and CO2) occurs only between blood capillaries and air capillaries. As air moves through a parabronchus and each successive air capillary, the partial pressure of oxygen (PO2) declines (as indicated by the decreased density of the stippling) because oxygen is diffusing into the blood capillaries associated with each air capillary. As a result of this diffusion, the partial pressure of oxygen in the blood leaving the lungs (pulmonary vein) is higher than that in blood entering the lungs (pulmonary artery) (as indicated by the increased density of the stippling).
Bottom: Relative partial pressures of O2 and CO2 (1) for air entering a parabronchus (initial-parabronchial,
PI) and air leaving a parabronchus (end-parabronchial, PE), and (2) for blood before entering blood capillaries in the lungs (pulmonary artery, PA) and for blood after leaving the blood capillaries in the lungs (pulmonary vein, PV). The partial pressure of oxygen (PO2) of
venous blood (PV) is derived from a mixture of all serial air capillary-blood
capillary
units. Because of this cross-current exchange the partial pressure of oxygen in avian pulmonary veins (PV) is greater than that of the air leaving the parabronchus (PE); air that will be exhaled. In mammals, the partial pressure of oxygen in veins leaving the lungs cannot
exceed
that of exhaled air (end-expiratory gas, or PE) (Figure adapted from Scheid and Piiper 1987).
Importantly, the partial pressure of oxygen in blood leaving the avian lung is the result of 'mixing'; blood from a series of capillaries associated with successive air capillaries along the length of a parabronchus is mixed as the blood leaves the capillaries and enters small veins. As a result, the direction of air flow through a parabronchus does not effect the efficiency of the cross-current exchange (because gases are only exchanged between blood capillaries and air capillaries, not between the parabronchus and the blood). So, in above diagram, reversing the direction of air flow would obviously mean that the air capillary on the far left would have the highest partial pressure of oxygen rather than the air capillary on the far right (so the stippling pattern that indicates the amount of oxygen in each air capillary would be reversed). However, because of the 'mixing' of blood just mentioned, this reversal would have little effect on the PV, the partial pressure of oxygen in blood leaving via pulmonary veins (the PO2 would likely be a bit lower because some oxygen would have been lost the first time air passed through the neopulmonic parabronchi). This is important because most birds have neopulmonic parabronchi as well as paleopulmonic parabronchi and, although air flow through paleopulmonic parabronchi is unidirectional, air flow through neopulmonic parabronchi is bidirectional.
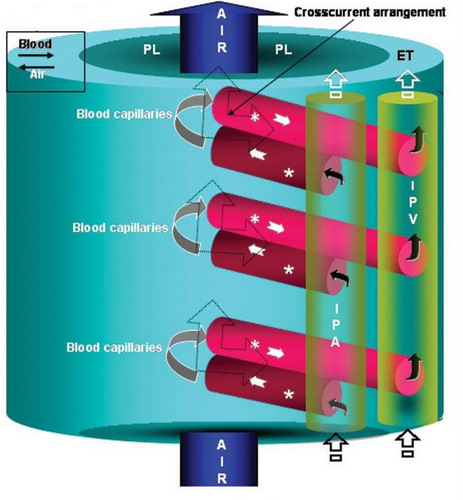
Diagram showing the flow of air from the parabronchial lumen (PL) into the air capillaries (not shown) and arterial blood from the periphery of the
parabronchus into the area of gas exchange (exchange tissue, ET). The orientation between the flow of air along the parabronchus and that
of blood into
the exchange tissue (ET) from the periphery is perpendicular or cross-current (dashed arrows). The exchange tissue is supplied with arterial blood
by interparabronchial arteries (IPA) that give rise to arterioles (stars) that terminate in blood capillaries. After passing through the capillaries, blood flows
into the intraparabronchial venules (asterisks) that drain into interparabronchial veins (IPV). These in turn empty into the pulmonary vein which returns the
blood to the heart. (From: Maina and Woodward 2009).
Control of Ventilation:
Ventilation and respiratory rate are regulated to meet the demands
imposed
by changes in metabolic activity (e.g., rest and flight) as well as
other
sensory inputs (e.g., heat and cold). There is likely a central
respiratory
control center in the avian brain, but this has not been unequivocally
demonstrated. As in mammals, the central control area appears to be
located
in the pons and medulla oblongata with facilitation and inhibition
coming
from higher regions of the brain. It also appears that the chemical
drive
on respiratory frequency and inspiratory and expiratory duration depend
on feedback from receptors in the lung as well as on extrapulmonary
chemoreceptors,
mechanoreceptors, and thermoreceptors (Ludders 2001).
Central chemoreceptors affect ventilation in response to changes in
arterial PCO2 and
hydrogen ion
concentration. Peripheral extrapulmonary chemoreceptors, specifically
the
carotid bodies (located in the carotid arteries), are influenced by PO2
and increase their discharge rate as PO2
decreases, thus increasing ventilation; they decrease their rate of
discharge
as PO2 increases or PCO2
decreases. These responses are the same as those observed in mammals.
Unlike
mammals, birds have a unique group of peripheral receptors located in
the
lung called intrapulmonary chemoreceptors (IPC) that are acutely
sensitive
to carbon dioxide and insensitive to hypoxia. The IPC affect rate and
volume
of breathing on a breath-to-breath basis by acting as the afferent limb
of an inspiratory-inhibitory reflex that is sensitive to the timing,
rate,
and extent of CO2 washout from the lung during
inspiration
(Ludders 2001).
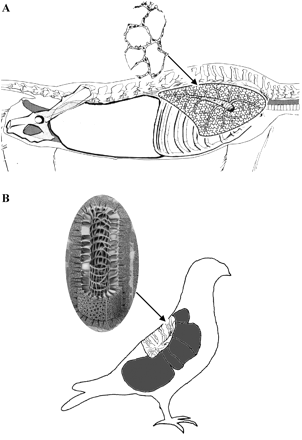
Respiration by Avian Embryos
During avian development there are
three sequential
stages of respiration (Tazawa 1987): prenatal (embryonic), paranatal
(hatching),
and postnatal (posthatching). During the prenatal stage respiratory gas
exchange occurs via diffusion between the external environment and the
initial gas exchanger (i.e., the area vasculosa, or the region of blood
island formation and forerunner of the chorioallantoic
membrane) in early embryonic life and later the vascular bed of the
chorioallantois. The paranatal stage starts when the beak penetrates
into
the air pocket (air cell) between the inner and outer shell membranes
(both
internal to shell; i.e., internal pipping) this occurs during the last
2-3 days of incubation. During this stage, the lungs begin to replace
the
chorioallantois as the gas exchanger, yet diffusion remains the major
mechanism
moving gas across the shell. The postnatal stage begins when the beak
penetrates
the shell (i.e., external pipping) (Brown et al. 1997).
Dubach, M. 1981. Quantitative analysis of the respiratory system of the House Sparrow, Budgerigar, and Violet-eared Hummingbird. Respiration Physiology 46: 43-60.
Duncker, H.-R. 1971. The lung air sac system of
birds.
Advances in Anatomy, Embryology, and Cell Biology 45: 1–171.
Hawkes, L. A. 2025. Why the superb physiological capacity of birds matters. Journal of Experimental Biology 228: jeb247986.
Klein, W., and T. Owerkowicz. 2006. Function of intracoelomic septa in lung ventilation of amniotes: lessons from lizards. Physiological and Biochemical Zoology 79: 1019-1032.
Ludders, J.W. 2001. Inhaled anesthesia for birds.
In: Recent advances in veterinary anesthesia and analgesia: companion
animals
(R. D. Gleed and J. W. Ludders, eds.). International Veterinary
Information
Service, Ithaca, NY. (www.ivis.org/advances/Anesthesia_Gleed/ludders2/chapter_frm.asp)
Maina, J.N. 1989. The morphometry of the avian lung.
Pp.
307-368 in Form and function in birds (A.S. King and J. McLelland,
eds.).
Academic Press, London.
Maina, J. N. 2002. Structure, function and evolution of the gas exchangers: comparative perspectives. Journal of Anatomy 201: 281-304.
Maina, J. N. 2008. Functional morphology of the avian respiratory system, the lung-air system: efficiency built on complexity. Ostrich 79: 117-132.
Maina, J. N., and A. S. King. 1989. The lung of the Emu, Dromaius novaehollandiae: a microscopic and morphometric study. Journal of Anatomy 163: 67-74.
Maina, J. N., A. S. King, and G. Settle. 1989. An allometric study of the pulmonary morphometric parameters in birds, with mammalian comparison. Philosophical Transactions of the Royal Society B 326: 1-57.
Maina, J. N., and J. D. Woodward. 2009. Three-dimensional serial section computer reconstruction of the arrangement of the structural components of the parabronchus of the Ostrich, Struthio camelus lung. Anatomical Record 292: 1685-1698.
McLelland, J. 1989. Anatomy of the lungs and air sacs. In: Form and function in birds, vol. 4 (A. S. King and J. McLelland, eds.), pp. 221-279. Academic Press, San Diego, CA.
Scheid, P., and J. Piiper. 1987. Gas exchange and transport. In: Bird respiration, volume 1 (T. J. Seller, ed.), pp. 97-129. CRC Press, Inc., Boca Raton, FL.
Sereno, P. C., R. N. Martinez, J. A. Wilson, D. J. Varricchio, O. A. Alcober, and H. C. E. Larsson. 2008. Evidence for avian intrathoracic air sacs in a new predatory dinosaur from Argentina. PLoS ONE 3: e3303.
Tazawa, H. 1987. Embryonic respiration. Pp. 3 - 24
in
Bird respiration, vol. 2 (T. J. Seller, ed.). CRC Press, Boca Raton, FL.
Tickle, P. G., A. R. Ennos, L. E. Lennox, S. F. Perry, and J. R. Codd. 2007. Functional significance of the uncinate processes in birds. Journal of Experimental Biology 210: 3955-3961.
Watson, R. R., Z. Fu, and J. B. West. 2007. Morphometry of the extremely thin pulmonary blood-gas barrier in the chicken lung. American Journal of Physiology. Lung Cellular and Molecular Physiology 36: L769-L777.
Wedel, M.J. 2003. Vertebral pneumaticity, air sacs,
and
the physiology of sauropod dinosaurs. Paleobiology 29: 243–255.
Welty, J.C. and L. Baptista. 1988. The life of
birds,
fourth edition. Saunders College Publishing, New York, NY.