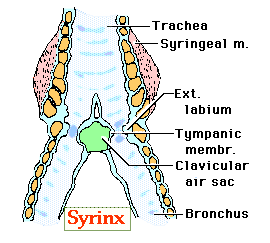
Source: http://trc.ucdavis.edu/mjguinan/apc100/modules/ Respiratory/airways/syrinx/syrinx.html |
 |
| BIO 554/754 Ornithology Vocal Communication |
|
Birds produce a variety sounds to communicate with flock members, mates (or potential mates), neighbors, & family members. These sounds vary from short, simple call notes (and short, simple songs like those of Henslow's Sparrows) . . .
. . . to surprisingly long, complex songs (e.g., the Superb Lyrebird).
Sometimes
birds generate sounds by using substrates (like woodpeckers)
or special feathers (like American
Woodcock) or special wings (like manakins).
Neat video of a Red-capped Manakin (Pipra mentalis) using its wings to generate sound.
Male Anna's Hummingbirds use their tail feathers to generate sound.
Most sounds, however, are produced by the avian vocal organ,
the
syrinx.
House Wren singing
Common Loon calling
|
Source: http://trc.ucdavis.edu/mjguinan/apc100/modules/ Respiratory/airways/syrinx/syrinx.html |
|
The syrinx is located at the point where the trachea branches into the two primary bronchi. According to one model of syrinx function, sound is generated when:
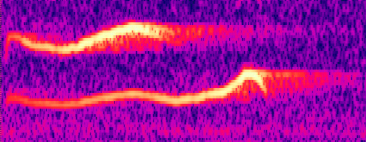
Click on the sonagram to hear a 'self-duet' by a Clay-colored
Robin
(Source: Doug Von Gausig's webpage at http://www.naturesongs.com/costa.html)
Wood Thrush
Characteristics of the sound (e.g., frequency) are influenced by vibrations of the internal tympaniform membrane (ITM). The 'characteristics' of the ITM (e.g., degree to which the membrane protrudes into the air pathway), in turn, are influenced by pressure in the interclavicular air sac (or, as in the diagram above, clavicular air sac) (Gill 1995). Syringeal muscles also influence air flow and the characteristics of sound (Gill 1995):
Superfast syringeal muscles -- Elemans et al. (2004) have found that Ring Doves (Streptopelia risoria) use "superfast" muscles to make their distinctive call. The dove's familiar cooing sound (see sonagram below) includes a trill, which is caused by an airflow that makes membranes in the syrinx vibrate. Those vibrations are made by activating two pairs of muscles which position the membranes in the airflow and tense or slacken them as appropriate to produce the trill. The only kind of muscles that can switch and on off as quickly as this are "superfast" ones, which are able to change direction in less than 10 microseconds. "Superfast" muscles have previously been identified in rattlesnakes, to help it agitate its rattle, and in the toadfish, a species of fish that makes foghorn-like noises to attract a mate. Given that doves have relatively simple calls, Elemans et al. (2004) believe that superfast vocal muscles may be quite widespread among birds, especially among songbirds. |
|

Spectrogram © 2004, Coen P.H. Elemans
The quality of sound can be further influenced by tracheal length, by constricting the larynx, by muscles in the throat, or by the structure and/or movements of the bill (e.g., here are some complex 'Bird Songs in Slow Motion').
Although the above model has been generally accepted for several
years,
Goller and Larsen (1997a, 1997b, 1999) provide evidence that other
structures
(not the ITM) are the source of sound in both songbirds (oscines) and
non-songbirds
because birds
can still vocalize when the medium (or internal) tympaniform membrane
is
experimentally kept from vibrating.
![]()

Each song syllable is accompanied by coordinated movements of the larynx and cornua that maintain an inverse relationship between
the size of the oropharyngeal–esophageal cavity and the song’s fundamental frequency. Left, Lateral view of cardinal showing the dorsoventral
movement (LV) of the larynx from the middle of the second cervical vertebra and its craniocaudal movement (LH) from the dorsal edge of the
beak-skull transition. Right, Ventrodorsal view showing distance between lateral cornua of hyoid apparatus (Cornua).
| Birdsong sounds sweeter because throats filter out messy overtones -- The purity of birdsong is owed in large part to rapid, controlled changes in the shape of the birds' upper vocal tracts. Riede et al. (2006) showed that songbirds adjust the size and shape of their vocal tract to 'fit' the changing frequency of their song, allowing birds to produce a more whistle-like, pure-tone song. These results support a growing consensus that birds and humans make sound in much the same way -- although it is presumed these processes evolved independently of each other in birds and hominids. Other studies have implicated tongues, beaks, especially beak gape, in shaping the sound that birds produce. Similarly, humans move their tongues, alter the shape of their upper vocal tracts, and change the shape of their mouths when they sing, laugh, talk and groan."The bird's vocal tract, like the human vocal tract in speech, acts as a resonance filter that can control the sound coming from the mouth," said co-author Rod Suthers. "Beak movements during song also contribute to this filter, but are not as important as changes in the size of the internal vocal tract. Human sopranos use the same technique as the cardinal to increase the loudness of very high notes so they can be heard above the orchestra."
That birds' throats vibrate when they sing will come as no surprise to birdwatchers. The effect of these oscillations on the birds' sound production, however, was unknown. The acoustics of sound-making are complicated. Most tones produced in nature are accompanied by a complex series of higher-pitched, quieter tones called overtones. When the loudness of these overtones is high, the tone sounds more complex. Birds can control the loudness of overtones to increase the tonal purity of their song. Humans use a similar technique to produce different vowel sounds of speech by altering the shapes of their throats, the positions of their tongues and the wideness of their mouths. "At low frequencies, the bird increases the volume of its oropharyngeal cavity and even expands the top of its esophagus," Suthers said. "These air-filled structures form a single cavity with a resonant frequency that matches the main frequency of the song. This amplifies the fundamental frequency and suppresses overtones." The shape and total volume (three-dimensional space, not loudness) of a singing Northern Cardinal's throat was observed and measured using X-ray cinematography to observe and measure the shape and total volume (three-dimensional space, not loudness) of a cardinal's throat as it spontaneously sang. Explanatory video can be viewed at: http://www.iuinfo.indiana.edu/bem/mr/rsfb/north_cardinal_large.mov (10 megs) The scientists determined that note changes in birdsong were accompanied by controlled changes in the volume of the upper esophagus as well as the positions of the bird's larynx and hyoid skeleton (a U-shaped bone formation in the bird's throat). They also found that the volume of the upper esophagus goes up whenever the main tone produced by the bird goes down, and vice versa. These alterations of shape have the effect of increasing the main tone and suppressing the loudness of overtones. |
![]()
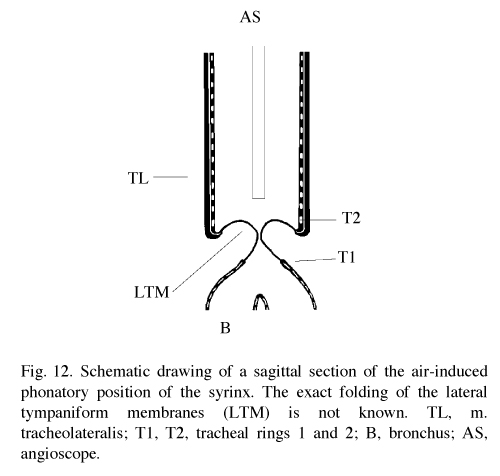 |
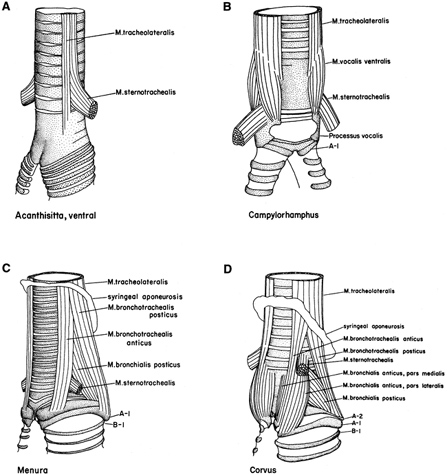
In pigeons (& other non-songbirds), there is a lateral (or external) tympaniform membrane (LTM) that spans two of the tracheal rings (T1 & T2) superior to the medial tympaniform membranes. It is these membranes that constrict the trachea, vibrate, and produce the sounds (Goller and Larsen 1997b). |
| In songbirds, "phonation is initiated by rostrad movement and stretching of the syrinx. At the same time, the syrinx is closed through movement of two soft tissue masses, the medial (ML) and lateral (LL) labia, into the bronchial lumen. Sound production always is accompanied by vibratory motions of both labia, indicating that these vibrations may be the sound source. However, because of the low temporal resolution of the imaging system, the frequency and phase of labial vibrations could not be assessed in relation to that of the generated sound. Nevertheless, in contrast to the previous model, these observations show that both labia contribute to aperture control and strongly suggest that they play an important role as principal sound generators" (Goller and Larson 1997a). | 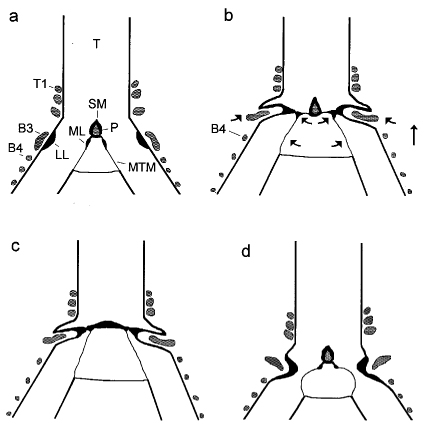  |
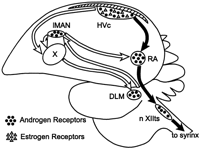
Central motor control of song:
Different circuits (or impulse pathways) in the brain control song production (posterior descending pathway) and song learning (anterior forebrain pathway). Song production is controlled via a pathway beginning in the brain & travelling to the syrinx:
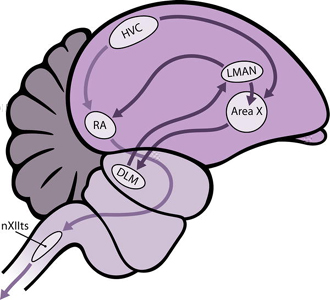
The 'learning pathway' connects the HVC to RA via areas X, DLM, and LMAN. This forms a recursive loop because the neurons in LMAN also project to area X. Disturbances to this pathway affect song development, but not the production of song in adult males.

| Virtual bird brain matches nature's tunes -- When birds sing, they force air from their lungs through the syrinx. Scientists at Rockefeller University and the University of Buenos Aires recently developed a simple computer model that mimics this process to produce sound. By simulating changes in the tension of the vocal folds and in the air pressure from the lungs, the model reproduced the song of a canary. But the song only sounded right if the lungs and the vocal folds vibrated with particular phase differences. How does a bird's brain give these complex commands? Some clues come from studies which show that a brain region called the high vocal centre is active when a bird sings. This activity excites neurons in the RA nucleus. Some neurons in this structure excite motor neurons that control muscles in the vocal folds or in the lungs. Others damp down the activity of nearby neurons. Laje et al. (2002) made a simple computer model of the RA neurons and were surprised to find that it changed a simple, constant signal from the high vocal centre into a complex series of bursts with the hallmark phase differences of birdsong. And when they fed the output signal from the virtual brain into their computer model of a bird's syrinx, it again sang like a bird. Simply varying the volume of the signal from the high vocal centre produced different song patterns. The model bird can accurately echo the song of the Chingolo sparrow (Zonotrichia capensis). The fake sparrow song sounds extremely similar to the real one. The big surprise is that the intricacies of birdsong arise from such simple instructions. Laje et al. plan to add more brainpower to the virtual bird, allowing it to listen as well as sing. This might help reveal how birds perfect their songs as they learn from other birds. -- Hazel Muir, New Scientist |
 Photo ©Mike Danzenbaker http://www.avesphoto.com/website/CH/species/SPARFC-1.htm |
Chingolo Sparrow singing
Testosterone (and melatonin; see below) appear to play some role in song production. For example:
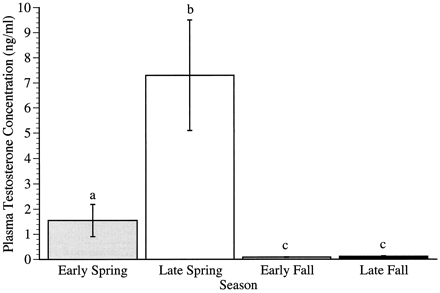
Seasonal changes in plasma testosterone concentrations
in male Song Sparrows.
Columns represent mean ± SEM (error bars) plasma
testosterone concentrations in male Song Sparrows
collected at each of the four sampling times (Smith et
al. 1997).

Male quality and owl hoots -- The evolution of communication through intrasexual selection is expected to lead signalers to transmit honest information on their fighting ability. Hardouin et al. (2007) studied the information encoded in the acoustic structure of the territorial calls of a nocturnal raptor. During territorial contests, male Scops Owls (Otus scops) give hoots composed of a downward frequency shift followed by a stable plateau. Hardouin et al. (2007) found that the frequency of the hoot was negatively correlated with the body weight of the vocalizer. They shifted the frequency of natural hoots to create resynthesized calls corresponding to individuals of varying body weight and used these stimuli in playback experiments simulating an intrusion into the territory of established breeders. Territory owners responded less intensely when they heard hoots simulating heavier intruders, and males with heavier apparent weight tended to give hoots with a lower frequency in response to playbacks simulating heavier intruders.
Although the current lack of understanding of the mechanisms of voice production in owls limits our ability to discuss the bases of this relationship, one possibility is that it may result from physiological constraints that operate during sound production. For example, lower-pitch hoots may be more costly to produce and/or reflect superior muscular or respiratory abilities. The relationship between pitch and body weight may reflect the fact that heavier, better-condition males are also characterized by higher testosterone levels, which in turn affect the frequency of their vocalizations. Indeed, male condition and testosterone levels have been shown to positively correlate, and higher testosterone levels are typically associated with more intense sexual displays. Moreover, experimental studies have demonstrated that injections of testosterone lower the frequency of male calls in birds, e.g., Gray Partridges (Perdrix perdrix) and Zebra Finches (Taeniopygia guttata).
Pygmy Owl (Glaucidium passerinum) hooting
| Melatonin Shapes Brain Structure In Songbirds -- Springtime's lengthening days spark the growth of gonads and a rush of sex hormones that drive songbirds to melodic flights of fancy. That much has been known for some time. But for the first time, Bentley et al. (1999) have also identified melatonin as a critical ingredient that regulates singing and fine-tunes the effects of testosterone on the brain."There is a lot of interest in melatonin," said co-author Gregory Ball, "but there has never been any indication that it affects brain plasticity like this. The fact that it would have a direct effect on a brain area in birds and influence its volume has never been addressed in other species." About 20 years ago, it was discovered that the high vocal center, or HVC, increases in volume as days grow longer. Scientists realized that longer daylight hours in the spring lead to a higher level of testosterone and prompts males to sing more. Scientists were able to link the rise in testosterone to physical changes in the HVC. But, scientists also noticed that even if songbirds are castrated, thus blocking the influence of testosterone, seasonal changes still affected the volume of HVC. "The changes weren't as large," observed Bentley, "but it was obvious that something else was controlling the change in volume. If it wasn't testosterone, what could it be? Because we knew that many hormones are controlled by photoperiod, we decided to look at an obvious candidate, melatonin." To conduct the study, European Starlings without testosterone were exposed to a range of artificial daylight hours that induced reproductive states characterized by different seasons. By providing birds with melatonin, researchers found they could still have a direct effect on the HVC, reducing overall volume or otherwise attenuating its growth despite the amount of daylight. "For example," Bentley said, "late in the summer, when birds terminate their reproductive activity and their gonads regress and testosterone disappears, the HVC doesn't really appear to shrink until later in the year. We think the effect of melatonin (which is secreted at night) is kicking in as the days get shorter, causing the volume of HVC to decrease slowly. To our knowledge, this is the first direct evidence of a role for melatonin in functional plasticity within the central nervous system of vertebrates." The next step is to find out how melatonin and testosterone interact to encourage efficiency in brain volumes during the breeding season. -- Source: John Hopkins University and Science Daily |
 www.mdc.missouri.gov
|
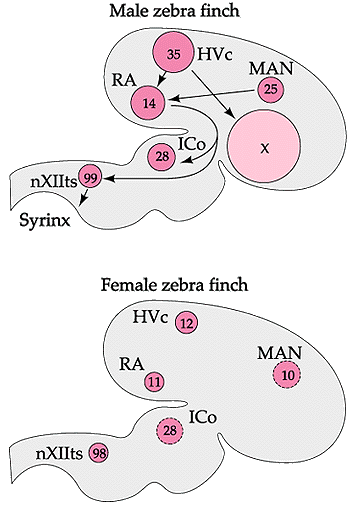
Sexual differentiation of the avian brain
In songbirds, males and females may have distinctly different brain
structures, specifically in those areas involved in the production of
song.
In many songbirds, males sing while females do not (or sing very
little).
The ability to sing is controlled by six different clusters of neurons
(nuclei) in the avian brain (see diagrams below). Neurons connect each
of these regions to one
another. In male songbirds, these nuclei can be several times larger
than the corresponding cluster of neurons in females, and in some
species
(e.g., Zebra Finches), females may lack one of these regions (area X)
entirely
(Arnold 1980, Konishi and Akutagawa 1985).
Classification of vocalizations:
Slaty-tailed Trogon calling
(Mayflower Bocawina National Park - Belize.. (more))
Nocturnal flight call of a Black-and-White Warbler
RealAudio
| AIFF
| WAV
(at 1/6 speed: RealAudio
| AIFF
| WAV)
Black-and-white Warbler at Mayflower Bocawina National Park, Belize ... (more)
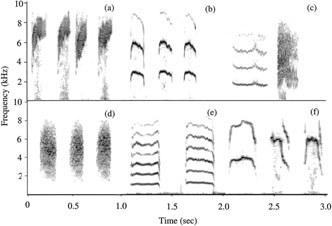 Sonograms of distress calls from six species. (a) Sooty- capped Bush-Tanager, (b) Black-capped Flycatcher, (c) Green Violet-ear (pictured below), (d) Gray-breasted Wood- Wren, (e) Streak-breasted Treehunter, and (f) Yellowish Flycatcher. Each sonogram represents 1 sec of distress calling.
|
Distress Calls of Birds in a Neotropical Cloud Forest (Neudorf and Sealy 2002 -- Distress calls are loud, harsh calls given by some species of birds when they are captured by a predator or handled by humans. Neudorf and Sealy (2002) recorded the frequency of distress calls in 40 species of birds captured in mist nets during the dry season in a Costa Rica cloud forest. They tested the following hypotheses proposed to explain the function of distress calls: (1) calling for help from kin or reciprocal altruists; (2) warning kin; (3) eliciting mobbing behavior; (4) startling the predator; and (5) distracting the predator through attraction of additional predators. Our results did not support the calling-for-help, warning kin, or mobbing hypotheses. Indeed, genera that regularly occurred with kin or in flocks were not more likely to call than non-flocking genera. There was no relationship between calling frequency and struggling behavior as predicted by the predator startle hypothesis. Larger birds tended to call more than smaller birds, providing some support for both the predator distraction hypothesis and predator startle hypotheses. Calls of higher amplitude may be more effective in startling the predator. Distress calls of larger birds may also travel greater distances than those of smaller birds, supporting the predator manipulation hypothesis. The adaptive significance of distress calls remains unclear as past studies have generated conflicting results. While more playback experiments are necessary to determine if calls indeed attract other individuals or predators, these results suggest that distress calls do not function to attract helpers or mobbers but are more likely directed toward predators. |
Kookaburra nest
Domestic Chicken - ground
predator alarm call
White-breasted Wood-wren giving alarm calls
(Mayflower Bocawina National Park - Belize)
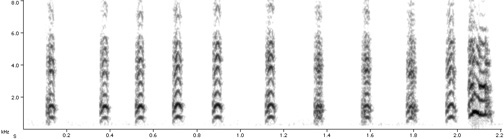
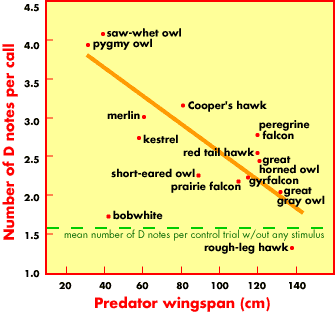 Predator wingspan compared to the number of "dee" tones on the end of the chickadees calls. The smaller (and more agile) the predator, the more "dees" get added, suggesting that chickadees recognize the danger of smaller predators. Hear a chickadee response to a Pygmy Owl - click here. Hear a chickadee response to a Great Horned Owl - click here. |
Chickadee language -- Black-capped
Chickadees
(Poecile atricapilla) have a complex language for warning
flock-mates
about predators. It was already known that chickadees utter a
high-pitched
"seet" when a predator was overhead, and used their "chick-a-dee" call
to, among other things, alert flock-mates to mob a threatening bird
that
was perched. However, Templeton et al. (2005) put flocks of six
chickadees
in an enclosure and recorded their responses. In the presence of a
harmless
quail, chickadees gave no alarm. But when a tethered raptor (hawk or
owl)
entered the cage, the alarms began. Alarms were more frequent when
Saw-whet
and Pygmy owls were present. But the alarms also had a different sound.
In the presence of small predators, the chickadees tacked an average of
four "dees" to their call: "chick-a-dee-dee-dee-dee." When the larger,
but less dangerous, Great Horned Owl was present, they used two dees:
"chick-a-dee-dee."
Smaller predators are more dangerous because of their greater agility
(see
graph to the left). To prove that the "language" was conveying information, Templeton et al. (2005) played back the recordings to chickadees. Recordings made in response to more dangerous raptors elicited more mobbing behavior, confirming that the chickadees understood the meaning of the calls. While this may be the most sophisticated bird "vocabulary" found to date, Templeton suspects others are out there. "This is the most detailed communication we have found, but it is also the finest scale that anyone has looked. It would not surprise me if other species are able to do this. All these signaling systems are a lot more complicated than we really expect, until we spend a lot of time and energy looking at them." -- David Tenenbaum |
Black-capped Chickadee
Nuthatches eavesdrop on chickadees -- Many animals recognize the alarm calls produced by other species, but the amount of information they glean from these eavesdropped signals is unknown. Black-capped Chickadees (Poecile atricapillus) have a sophisticated alarm call system in which they encode complex information about the size and risk of potential predators in variations of a single type of mobbing alarm call. Templeton and Greene (2007) showed experimentally that Red-breasted Nuthatches (Sitta canadensis) respond appropriately to variation in heterospecific "chick-a-dee" alarm calls (i.e., stronger mobbing behavior to playback of small-predator alarm calls), indicating that they gain important information about potential predators in their environment. These results demonstrate a previously unsuspected level of discrimination in intertaxon eavesdropping.

Siberian Jay (Photo by John van der Woude)
Calls 'describe' predator's behavior -- Predation is a powerful agent of natural selection, driving the evolution of antipredator calls. These calls have been shown to communicate predator category and/or predator distance to conspecifics. However, the risk posed by predators depends also on predator behavior, and the ability of prey to communicate predator behavior to conspecifics would be a selective advantage reducing their predation risk. Griesser (2008) tested this idea within Siberian Jays (Perisoreus infaustus), a group-living bird species. Predation by hawks, and to a lesser extent by owls, is substantial and the sole cause of mortality in adult jays. Field data and predator-exposure experiments revealed that jays used antipredator calls for hawks depending on predator behavior. A playback experiment demonstrated that these prey-to-prey calls were specific to hawk behavior (perch, prey search, or attack) and elicited distinct, situation-specific escape responses. This is the first study to demonstrate that prey signals convey information about predator behavior to conspecifics. Given that antipredator calls by jays serve to protect kin group members, consequently lowering their mortality, kin-selected benefits could be an important factor for the evolution of predator-behavior-specific antipredator calls in such systems.
| Low frequency calls of cassowaries --
Although
some birds can detect wavelengths in the infrasound range,there has
been
litle evidence that birds produce very low frequencies. Mack and Jones
(2003) made 9 recordings of a captive Dwarf Cassowary (Casuarius
benneti)
and one recording of a wild Southern Cassowary (C. casuarius)
in Papua New Guinea. Both species produced sounds near the floor of the
human hearing range in their pulsed booming notes: down to 32 Hz for C.
casuarius
and 23 Hz in C. benneti. Natural selection should favor the evolution of vocalizations that reach their targets with minimal degradation, and low frequencies propagate over long distances with minimal attenuation caused by vegetation. New Guinea forests ofen have a fairly thick understory of wet leafy vegetation that could quickly attenuate higher frequencies. Thus, the very low frequency calls of cassowaries probably ideal for communication among widely dispersed, solitary cassowaries in dense rainforest. How cassowaries produce such low vocalizations is currently unknown. All three cassowary species have keratinous casques rising from the upper mandible over the top of the skull up to 17 cm in height. Hypotheses concerning the function of the casque include: (1) a secondary sexual character, (2) a weapon in dominance disputes, (3) a tool for scraping the leaf-litter, or (4) a crash helmet for birds as they bash through the undergrowth. The later three seem unlikely based on field observations. Future research should include the possibility that the casque might play some role in sound reception or acoustic communication. |
 Photo by D. DeMello, Wildlife Conservation Society |
The functions of bird song may vary among species. Some known & hypothesized functions include:

Birds Reveal their Personality when Singing -- Individual differences in social behavior may have consequences for mate choice and sexual signalling, because partners should develop preferences for personalities that maximize reproductive output. Here we propose that behavioural traits involved in sexual advertisement may serve as good indicators of personality, which is fundamental for sexual selection to operate on temperament. Bird song has a prominent and well-established role in sexual selection, and it displays considerable variation among individuals with a potentially strong personality component.
In a field study of free-living male Collared Flycatchers (Ficedula albicollis), Garamszegi et al. (2008) characterized personality based on the exploration of an altered breeding environment, and based on the risk taken when a potential predator was approaching during a simulated territorial interaction. They found that explorative and risk-taker individuals consistently sang at lower song posts than shy individuals in the presence of a human observer. Moreover, males from lower posts established pair-bonds relatively faster than males from higher posts.
These results may demonstrate that risk taking during singing correlates with risk taking during aggression and with exploration, thus personality may be manifested in different contexts involving sexual advertisement. These findings are in accordance with the hypothesis that the male's balance between investment in reproduction and risk taking is reflected in sexual displays, and it may be important information for choosy females that seek partners with personality traits enhancing breeding success.