Birds produce a variety sounds to communicate with flock members, mates (or potential mates), neighbors, & family members. These sounds vary from short, simple call notes (and short, simple songs like those of Henslow's Sparrows) . . .
Henslow's Sparrow
. . . to surprisingly long, complex songs (e.g., the Superb Lyrebird).
Superb Lyrebird
Sometimes birds generate sounds by using substrates (like woodpeckers) or their wings (like Ruffed Grouse) or special feathers (like American Woodcock) or special wings (like manakins).
Most woodpeckers communicate by hammering their bill repeatedly against a tree, which produces an acoustic signal used in courtship and competition. This loud atonal signal is called drumming. Each time the bill makes contact with the tree it produces a drum beat. Ornithologists have long known that drumming is a territorial signal produced by resident birds as they settle and defend their territories
during the breeding season. Drumming as a social
signal that can function over both short and long distances. During agonistic encounters, for example, individuals
perform drums when opponents invade their territories. In addition, residents often drum at specific times in
the day (e.g., dawn), much like songbirds that sing at morning's first
light to advertise to neighbors that they still occupy the
area.
(A) During the breeding season, Downy Woodpeckers excavate nest cavities
in trees using their bill. (B) Woodpeckers also use their bill to forage on trees for food, including probing for insects. (C) Most woodpeckers
communicate by hammering their bill repeatedly against a tree, producing an acoustic signal (drumming) used in courtship and competition.
Studies of several woodpecker species have revealed that playback of drums over speakers (to simulate terrritorial intrusion by a conspecific) elicits aggressive behavior by the resident birds, including both males and females. For example, in response to playback of drums, male and female Downy Woodpeckers engage in a wide range of behaviors to defend their territories, including calls and attack flights, as well as drums. Recent experimental work has revealed that woodpeckers may vary drum speed (i.e., varying the duration between successive beats in a drum), duration, and rhythm during territorial contests. For example, Red-bellied Woodpeckers actively engaged in territorial interactions produce drums longer in duration than drums produced spontaneously in
the morning. Woodpeckers responding to conspecifics in territorial disputes may also produce drums with shorter inter-beat intervals Thus, woodpeckers can adjust how they
drum to regulate the effectiveness or potency of the signal in a
context-dependent manner.
Drumming is a high-speed activity. A plot showing the average drumming rates of different species of woodpeckers (organized by genus) reveals that (i) drumming speeds vary both within and across genera; and (ii) many
species drum at rates exceeding 20 beats per second (Figure below). Speed matters because fast motions place demands on the neuromuscular system, and thus the evolution of fast drumming may have been driven by its usefulness as an honest signal of condition.
Variability in drumming frequency (beats per second) within and between
22 woodpecker genera. Each point represents the mean frequency of
a single species (Text and figures from: Schuppe et al. 2021).
Downy Woodpecker drumming
Most sounds, however, are produced by the avian vocal organ, the syrinx.
The syrinx is located at the point where the trachea branches into the two primary bronchi. According to one model of syrinx function, sound is generated when:
contraction of muscles (thoracic & abdominal) force air from air sacs through the bronchi & syrinx
the air molecules vibrate as they pass through the narrow passageways between the external labia & the internal tympaniform membranes (or, as in the diagram above, tympanic membrane.
With two separate passageways (and membranes), some birds are able to generate two different sounds at the same time:
The ultimate trill. The Wood Thrush song ends with one of the most complex sounds a bird can create. Layered above a series of lower-pitch mini-trills is a series of higher-pitch sweeping tones. To perform this impressive trill the bird must pair impeccably timed breath control with independent muscle movements for each side of the syrinx.
Characteristics of the sound (e.g., frequency) are influenced by vibrations of the internal tympaniform membrane (ITM). The 'characteristics' of the ITM (e.g., degree to which the membrane protrudes into the air pathway), in turn, are influenced by pressure in the interclavicular air sac (or, as in the diagram above, clavicular air sac) (Gill 1995). Syringeal muscles also influence air flow and the characteristics of sound (Gill 1995):
most nonpasserines have 2 pairs of muscles on the sides of the trachea above & outside the syrinx (extrinsic muscles)
passerines have extrinsic muscles plus intrinsic syringeal muscles (up to 6 pairs) that originate in the syrinx & insert on bronchial rings, internal & external tympaniform membranes & syringeal cartilage
Vocal organs of passerine birds. Syringes (vocal organs) of representative passerines are located
at the juncture of the trachea and bronchi. (a) Acanthisitta (New Zealand wren) showing a relatively
simple syrinx lacking complex musculature; (b) Campylorhamphus (woodcreeper) illustrating
a tracheophone syrinx; (c) Menura (lyrebird) showing an atypical oscine syrinx;
(d) Corvus (crow) showing a typical oscine syrinx. (Raikow and Bledsoe 2000). The avian syrinx
is categorized based on its location: 1. Tracheophone or Tracheal = in trachea (most New World suboscine
Furnari Passeriformes), 2. Haploophone or Bronchial = in bronchi (cuckoos, nightjars), and
3. Tracheobronchial = at junction of trachea and bronchi (oscine Passeriformes).
Superfast syringeal muscles -- Elemans et al. (2004) have found that Ring Doves (Streptopelia risoria) use "superfast" muscles to make their distinctive call. The dove's familiar cooing sound (see sonagram below) includes a trill, which is caused by an airflow that makes membranes in the syrinx vibrate. Those vibrations are made by activating two pairs of muscles which position the membranes in the airflow and tense or slacken them as appropriate to produce the trill. The only kind of muscles that can switch and on off as quickly as this are "superfast" ones, which are able to change direction in less than 10 microseconds. "Superfast" muscles have previously been identified in rattlesnakes, to help it agitate its rattle, and in the toadfish, a species of fish that makes foghorn-like noises to attract a mate. Given that doves have relatively simple calls, Elemans et al. (2004) believe that superfast vocal muscles may be quite widespread among birds, especially among songbirds.
The quality of sound can be further influenced by tracheal length, by constricting the larynx, by muscles in the throat, or by the structure and/or movements of the bill (e.g., here are some complex 'Bird Songs in Slow Motion').
Although the above model has been generally accepted for several years, Goller and Larsen (1997a, 1997b, 1999) provide evidence that other structures (not the ITM) are the source of sound in both songbirds (oscines) and non-songbirds because birds can still vocalize when the medium (or internal) tympaniform membrane is experimentally kept from vibrating.
Each song syllable is accompanied by coordinated movements of the larynx and cornua that maintain an inverse relationship between
the size of the oropharyngeal–esophageal cavity and the song’s fundamental frequency. Left, Lateral view of cardinal showing the dorsoventral
movement (LV) of the larynx from the middle of the second cervical vertebra and its craniocaudal movement (LH) from the dorsal edge of the
beak-skull transition. Right, Ventrodorsal view showing distance between lateral cornua of hyoid apparatus (Cornua).
Birdsong sounds sweeter because throats filter out messy overtones-- The purity of birdsong is owed in large part to rapid, controlled changes in the shape of the birds' upper vocal tracts. Riede et al. (2006) showed that songbirds adjust the size and shape of their vocal tract to 'fit' the changing frequency of their song, allowing birds to produce a more whistle-like, pure-tone song. These results support a growing consensus that birds and humans make sound in much the same way -- although it is presumed these processes evolved independently of each other in birds and hominids. Other studies have implicated tongues, beaks, especially beak gape, in shaping the sound that birds produce. Similarly, humans move their tongues, alter the shape of their upper vocal tracts, and change the shape of their mouths when they sing, laugh, talk and groan."The bird's vocal tract, like the human vocal tract in speech, acts as a resonance filter that can control the sound coming from the mouth," said co-author Rod Suthers. "Beak movements during song also contribute to this filter, but are not as important as changes in the size of the internal vocal tract. Human sopranos use the same technique as the cardinal to increase the loudness of very high notes so they can be heard above the orchestra."
That birds' throats vibrate when they sing will come as no surprise to birdwatchers. The effect of these oscillations on the birds' sound production, however, was unknown. The acoustics of sound-making are complicated. Most tones produced in nature are accompanied by a complex series of higher-pitched, quieter tones called overtones. When the loudness of these overtones is high, the tone sounds more complex. Birds can control the loudness of overtones to increase the tonal purity of their song. Humans use a similar technique to produce different vowel sounds of speech by altering the shapes of their throats, the positions of their tongues and the wideness of their mouths.
"At low frequencies, the bird increases the volume of its oropharyngeal cavity and even expands the top of its esophagus," Suthers said. "These air-filled structures form a single cavity with a resonant frequency that matches the main frequency of the song. This amplifies the fundamental frequency and suppresses overtones." The shape and total volume (three-dimensional space, not loudness) of a singing Northern Cardinal's throat was observed and measured using X-ray cinematography to observe and measure the shape and total volume (three-dimensional space, not loudness) of a cardinal's throat as it spontaneously sang.
The scientists determined that note changes in birdsong were accompanied by controlled changes in the volume of the upper esophagus as well as the positions of the bird's larynx and hyoid skeleton (a U-shaped bone formation in the bird's throat). They also found that the volume of the upper esophagus goes up whenever the main tone produced by the bird goes down, and vice versa. These alterations of shape have the effect of increasing the main tone and suppressing the loudness of overtones.
In pigeons (& other non-songbirds), there is a lateral (or external) tympaniform membrane (LTM) that spans two of the tracheal rings (T1 & T2) superior to the medial tympaniform membranes. It is these membranes that constrict the trachea, vibrate, and produce the sounds (Goller and Larsen 1997b).
Wood Thrush using both sides of the syrinx in a single song
In songbirds, "phonation is initiated by rostrad movement and stretching of the syrinx. At the same time, the syrinx is closed through movement of two soft tissue masses, the medial (ML) and lateral (LL) labia, into the bronchial lumen. Sound production always is accompanied by vibratory motions of both labia, indicating that these vibrations may be the sound source. However, because of the low temporal resolution of the imaging system, the frequency and phase of labial vibrations could not be assessed in relation to that of the generated sound. Nevertheless, in contrast to the previous model, these observations show that both labia contribute to aperture control and strongly suggest that they play an important role as principal sound generators" (Goller and Larson 1997a).
Central motor control of song:
Different circuits (or impulse pathways) in the brain control song production (posterior descending pathway) and song learning (anterior forebrain pathway). Song production is controlled via a pathway beginning in the brain & travelling to the syrinx:
Impulses start in the HVC (High Vocal Center in the neostriatum), pass to the RA (Robust nucleus of the Archistriatum), then to the tracheosyringeal motor (hypoglossal) nucleus (nXIIts) in the brain stem, and finally along motor neurons to the syringeal muscles.
The acquisition and production of learned song is made possible by a group of discrete brain nuclei and their connecting pathways, referred to as the “song system,” which has similarities in the three groups of birds—songbirds, parrots, and hummingbirds—that evolved learned song. This system has two main branches: the posterior descending pathway (PDP), necessary for both the acquisition and production of learned song, and the anterior forebrain pathway (AFP), necessary for acquisition only. The high vocal center (HVC) is at the starting point of both these pathways, but the HVC cells that project to the PDP and AFP differ. In mammalian terms the PDP is homologous to a motor pathway that starts in the cerebral cortex and descends through the brain stem, while the AFP is homologous to a cortical pathway through the basal ganglia and thalamus (Nottebohm 2005).
The 'learning pathway' connects the HVC to RA via areas X, DLM, and LMAN. This forms a recursive loop because the neurons in LMAN also project to area X. Disturbances to this pathway affect song development, but not the production of song in adult males.
The neurobiology of birdsong
Schematic diagrams of parasagittal views of the brains of a songbird (a) and a non-songbird (b) illustrate the dramatic differences between them. Songbirds have an elaborate network of interconnected forebrain nuclei that form an interface between auditory input (which converges on field L, the primary auditory projection region in the avian forebrain) and vocal output generated by the syrinx, the avian vocal organ. Non-songbirds also have field L, and they can produce vocalizations in the syrinx, but they do not have the network of forebrain nuclei that songbirds have. DLM, nucleus dorsolateralis anterior, pars medialis; DM, dorsomedial nucleus of the midbrain nucleus intercollicularis; HVC, a letter based name; lMAN, lateral magnocellular nucleus of the anterior nidopallium; mMAN, medial magnocellular nucleus of the anterior nidopallium; NIF, nucleus interface of the nidopallium; nXIIts, tracheosyringeal portion of the nucleus hypoglossus; RA, robust nucleus of the arcopallium; RAm, nucleus retroambigualis; rVRG, rostro–ventral respiratory group; X, Area X (Bolhuis and Gahr 2006).
Virtual bird brain matches nature's tunes -- When birds sing, they force air from their lungs through the syrinx. Scientists at Rockefeller University and the University of Buenos Aires recently developed a simple computer model that mimics this process to produce sound. By simulating changes in the tension of the vocal folds and in the air pressure from the lungs, the model reproduced the song of a canary. But the song only sounded right if the lungs and the vocal folds vibrated with particular phase differences. How does a bird's brain give these complex commands? Some clues come from studies which show that a brain region called the high vocal centre is active when a bird sings. This activity excites neurons in the RA nucleus. Some neurons in this structure excite motor neurons that control muscles in the vocal folds or in the lungs. Others damp down the activity of nearby neurons. Laje et al. (2002) made a simple computer model of the RA neurons and were surprised to find that it changed a simple, constant signal from the high vocal centre into a complex series of bursts with the hallmark phase differences of birdsong. And when they fed the output signal from the virtual brain into their computer model of a bird's syrinx, it again sang like a bird. Simply varying the volume of the signal from the high vocal centre produced different song patterns. The model bird can accurately echo the song of the Rufous-collared Sparrow (Zonotrichia capensis). The fake sparrow song sounds extremely similar to the real one. The big surprise is that the intricacies of birdsong arise from such simple instructions. Laje et al. plan to add more brainpower to the virtual bird, allowing it to listen as well as sing. This might help reveal how birds perfect their songs as they learn from other birds. -- Hazel Muir, New Scientist
Testosterone (and melatonin; see below)appear to play some role in song production. For example:
Autoradiographic studies have shown that the neurons of the song-controlling nuclei incorporate radioactive testosterone, whereas other regions of the brain do not (Arnold et al. 1976).
Male Zebra Finches - correlation between the amount of song & the concentration of serum testosterone (Pröve 1978)
Seasonal changes in testosterone levels are correlated with the seasonal singing patterns
When testosterone levels are low, there is a decrease in song & a decrease in the size of the male-specific brain nuclei (Nottebohm 1981).
In adult Chaffinches, castration eliminates song, but injection of testosterone induces such birds to sing even in November, when they are normally silent (Thorpe 1958).
Females in some species can be induced to sing by injecting them with testosterone (Nottebohm 1980).
Seasonal changes in plasma testosterone concentrations in male Song Sparrows.
Columns represent mean ± SEM (error bars) plasma testosterone concentrations in male Song Sparrows
collected at each of the four sampling times (Smith et al. 1997).
Spectrograms of hoots from three different males showing the variation in frequency.
Male quality and owl hoots -- The evolution of communication through intrasexual selection is expected to lead signalers to transmit honest information on their fighting ability. Hardouin et al. (2007) studied the information encoded in the acoustic structure of the territorial calls of a nocturnal raptor. During territorial contests, male Scops Owls (Otus scops) give hoots composed of a downward frequency shift followed by a stable plateau. Hardouin et al. (2007) found that the frequency of the hoot was negatively correlated with the body weight of the vocalizer. They shifted the frequency of natural hoots to create resynthesized calls corresponding to individuals of varying body weight and used these stimuli in playback experiments simulating an intrusion into the territory of established breeders. Territory owners responded less intensely when they heard hoots simulating heavier intruders, and males with heavier apparent weight tended to give hoots with a lower frequency in response to playbacks simulating heavier intruders.
Although the current lack of understanding of the mechanisms of voice production in owls limits our ability to discuss the bases of this relationship, one possibility is that it may result from physiological constraints that operate during sound production. For example, lower-pitch hoots may be more costly to produce and/or reflect superior muscular or respiratory abilities. The relationship between pitch and body weight may reflect the fact that heavier, better-condition males are also characterized by higher testosterone levels, which in turn affect the frequency of their vocalizations. Indeed, male condition and testosterone levels have been shown to positively correlate, and higher testosterone levels are typically associated with more intense sexual displays. Moreover, experimental studies have demonstrated that injections of testosterone lower the frequency of male calls in birds, e.g., Gray Partridges (Perdrix perdrix) and Zebra Finches (Taeniopygia guttata).
Melatonin Shapes Brain Structure In Songbirds -- Springtime's lengthening days spark the growth of gonads and a rush of sex hormones that drive songbirds to melodic flights of fancy. That much has been known for some time. But for the first time, Bentley et al. (1999) have also identified melatonin as a critical ingredient that regulates singing and fine-tunes the effects of testosterone on the brain."There is a lot of interest in melatonin," said co-author Gregory Ball, "but there has never been any indication that it affects brain plasticity like this. The fact that it would have a direct effect on a brain area in birds and influence its volume has never been addressed in other species." About 20 years ago, it was discovered that the high vocal center, or HVC, increases in volume as days grow longer. Scientists realized that longer daylight hours in the spring lead to a higher level of testosterone and prompts males to sing more. Scientists were able to link the rise in testosterone to physical changes in the HVC. But, scientists also noticed that even if songbirds are castrated, thus blocking the influence of testosterone, seasonal changes still affected the volume of HVC. "The changes weren't as large," observed Bentley, "but it was obvious that something else was controlling the change in volume. If it wasn't testosterone, what could it be? Because we knew that many hormones are controlled by photoperiod, we decided to look at an obvious candidate, melatonin." To conduct the study, European Starlings without testosterone were exposed to a range of artificial daylight hours that induced reproductive states characterized by different seasons. By providing birds with melatonin, researchers found they could still have a direct effect on the HVC, reducing overall volume or otherwise attenuating its growth despite the amount of daylight. "For example," Bentley said, "late in the summer, when birds terminate their reproductive activity and their gonads regress and testosterone disappears, the HVC doesn't really appear to shrink until later in the year. We think the effect of melatonin (which is secreted at night) is kicking in as the days get shorter, causing the volume of HVC to decrease slowly. To our knowledge, this is the first direct evidence of a role for melatonin in functional plasticity within the central nervous system of vertebrates." The next step is to find out how melatonin and testosterone interact to encourage efficiency in brain volumes during the breeding season. -- Source: John Hopkins University and Science Daily
Simplified view of the avian song control system showing distribution
of steroid receptors. HVc, High
Vocal Center (Brenowitz and
Lent 2002).
Sexual differentiation of the avian brain
In songbirds, males and females may have distinctly different brain structures, specifically in those areas involved in the production of song. In many songbirds, males sing while females do not (or sing very little). The ability to sing is controlled by six different clusters of neurons (nuclei) in the avian brain (see diagrams below). Neurons connect each of these regions to one
another. In male songbirds, these nuclei can be several times larger than the corresponding cluster of neurons in females, and in some species (e.g., Zebra Finches), females may lack one of these regions (area X) entirely (Arnold 1980, Konishi and Akutagawa 1985).
Classification of vocalizations:
Songs
primarily under the influence of sex hormones
generally important in reproduction (e.g., defending territories & attracting mates)
Calls
generally concerned with coordination of the behavior of a pair, family group, or flock
not primarily sexual, but important in 'maintenance' activities, such as foraging, flocking, & responding to threats of predation
usually are acoustically simple
may serve a variety of functions:
location/contact/individual recognition
Elegant Trogon calling
nocturnal flight calls. These calls help birds form and maintain in-flight associations, and also provide locational information that helps flying birds avoid collisions.
distress call - uttered when captured by a predator (or when being removed from a mist-net by an ornithologist)
Pileated Woodpecker distress calls
Sonograms of distress calls of six species. (a) Sooty-capped Bush-Tanager, (b) Black-capped Flycatcher, (c) Green Violet-ear, (d) Gray-breasted Wood-Wren, (e) Streak-breasted Treehunter, and (f) Yellowish Flycatcher. Each sonogram represents 1 sec of calling.
Distress Calls of Birds in a Neotropical Cloud Forest (Neudorf and Sealy 2002 -- Distress calls are loud, harsh calls given by some species of birds when they are captured by a predator or handled by humans. Neudorf and Sealy (2002) recorded the frequency of distress calls in 40 species of birds captured in mist nets during the dry season in a Costa Rica cloud forest. They tested the following hypotheses proposed to explain the function of distress calls: (1) calling for help from kin or reciprocal altruists; (2) warning kin; (3) eliciting mobbing behavior; (4) startling the predator; and (5) distracting the predator through attraction of additional predators. Our results did not support the calling-for-help, warning kin, or mobbing hypotheses. Indeed, genera that regularly occurred with kin or in flocks were not more likely to call than non-flocking genera. There was no relationship between calling frequency and struggling behavior as predicted by the predator startle hypothesis. Larger birds tended to call more than smaller birds, providing some support for both the predator distraction hypothesis and predator startle hypotheses. Calls of higher amplitude may be more effective in startling the predator. Distress calls of larger birds may also travel greater distances than those of smaller birds, supporting the predator manipulation hypothesis. The adaptive significance of distress calls remains unclear as past studies have generated conflicting results. While more playback experiments are necessary to determine if calls indeed attract other individuals or predators, these results suggest that distress calls do not function to attract helpers or mobbers but are more likely directed toward predators.
Scroll up to about the 0:47 mark to hear a European Starling give distress calls while being removed from a mist-net.
feeding
aggression
courtship
copulation or post-copulatory
begging
Kookaburra nest
alarm (aerial predator vs. ground predator)
Domestic Chicken - aerial predator alarm call
Domestic Chicken - ground predator alarm call
Alarm calls
Predator wingspan compared to the number of "dee" tones
on the end of the chickadees calls. The smaller (and more
agile) the predator, the more "dees" get added, suggesting
that chickadees recognize the danger of smaller predators.
Hear a chickadee response to a Pygmy Owl - click here.
Hear a chickadee response to a Great Horned Owl - click here.
Chickadee language -- Black-capped Chickadees (Poecile atricapilla) have a complex language for warning flock-mates about predators. It was already known that chickadees utter a high-pitched "seet" when a predator was overhead, and used their "chick-a-dee" call to, among other things, alert flock-mates to mob a threatening bird that was perched. However, Templeton et al. (2005) put flocks of six chickadees in an enclosure and recorded their responses. In the presence of a harmless quail, chickadees gave no alarm. But when a tethered raptor (hawk or owl) entered the cage, the alarms began. Alarms were more frequent when Saw-whet and Pygmy owls were present. But the alarms also had a different sound. In the presence of small predators, the chickadees tacked an average of four "dees" to their call: "chick-a-dee-dee-dee-dee." When the larger, but less dangerous, Great Horned Owl was present, they used two dees: "chick-a-dee-dee." Smaller predators are more dangerous because of their greater agility (see graph to the left). To prove that the "language" was conveying information, Templeton et al. (2005) played back the recordings to chickadees. Recordings made in response to more dangerous raptors elicited more mobbing behavior, confirming that the chickadees understood the meaning of the calls. While this may be the most sophisticated bird "vocabulary" found to date, Templeton suspects others are out there. "This is the most detailed communication we have found, but it is also the finest scale that anyone has looked. It would not surprise me if other species are able to do this. All these signaling systems are a lot more complicated than we really expect, until we spend a lot of time and energy looking at them." -- David Tenenbaum
Chris Templeton: "How Chickadees Communicate about Predators
Black-capped Chickadee responding to playback of recording of an Eastern Screech-Owl
Nuthatches eavesdrop on chickadees -- Many animals recognize the alarm calls produced by other species, but the amount of informationthey glean from these eavesdropped signals is unknown. Black-capped Chickadees (Poecile atricapillus) havea sophisticated alarm call system in which they encode complexinformation about the size and risk of potential predators in variations of a single type of mobbing alarm call. Templeton and Greene (2007) showed experimentally that Red-breasted Nuthatches (Sitta canadensis) respond appropriately to variation in heterospecific"chick-a-dee" alarm calls (i.e., stronger mobbing behavior to playback of small-predator alarm calls), indicating that they gain important information about potential predators in theirenvironment. These results demonstrate a previously unsuspectedlevel of discrimination in intertaxon eavesdropping.
Calls 'describe' predator's behavior -- Predation is a powerful agent of natural selection, driving the evolution of antipredator calls. These calls have been shown to communicate predator category and/or predator distance to conspecifics. However, the risk posed by predators depends also on predator behavior, and the ability of prey to communicate predator behavior to conspecifics would be a selective advantage reducing their predation risk. Griesser (2008) tested this idea within Siberian Jays (Perisoreus infaustus), a group-living bird species. Predation by hawks, and to a lesser extent by owls, is substantial and the sole cause of mortality in adult jays. Field data and predator-exposure experiments revealed that jays used antipredator calls for hawks depending on predator behavior. A playback experiment demonstrated that these prey-to-prey calls were specific to hawk behavior (perch, prey search, or attack) and elicited distinct, situation-specific escape responses. This is the first study to demonstrate that prey signals convey information about predator behavior to conspecifics. Given that antipredator calls by jays serve to protect kin group members, consequently lowering their mortality, kin-selected benefits could be an important factor for the evolution of predator-behavior-specific antipredator calls in such systems.
Sound spectrograms for two types of Great Tit alarm calls: (A) a ‘chicka’ alarm call consisting of three distinct types of syllables and (B) a ‘jar’ alarm call consisting of a single type of syllables that vary in duration. (C and D) Number of alarm calls (mean ± SE) given by Great Tit parents in the first 5 minutes in response to presentation of a jungle crow (n = 11) or a Japanese rat snake (n = 10) near their nest, (C) ‘chicka’ alarm calls, and (D) ‘jar’ alarm calls. (E and F) Nestling responses to parental alarm calls given for a crow (n = 11) and for a snake (n = 10). (E) Percentage of trials where the amount of time nestlings spent crouching motionless increased after parents gave alarm calls, and (F) percentage of trials where nestlings jumped out of the nest box in response to alarm calls.
Alarm calls warn nestlings of different threats -- Animal communication signals can contain surprisingly complex information, which plays a vital role in a variety of social interactions. For example, many species of birds and mammals produce vocal alarm signals when encountering a predator, and these calls often serve to communicate the type of predator and/or the degree of danger to members of a social group. Similarly, signals used in parent–offspring interactions can encode sophisticated information such as the type and immediacy of threat to the offspring. Suzuki (2011) found that differential use of parental alarm calls in Great Tits (Parus major) functions to elicit different predator-avoidance behaviors in altricial nestlings: great tit parents produce acoustically distinctive alarm calls for the two main nest predators, the Jungle Crow (Corvus macrorhynchos) and the Japanese rat snake (Elaphe climacophora). Nestlings crouched down inside their nest cavity in response to alarm calls given for a crow, whereas they fled the cavity in response to alarm calls given for a snake. The two responses help nestlings to selectively evade those predators because crows snatch nestlings from the nest entrance, whereas snakes invade the nest cavity. Although chicks of some species have been shown to recognize and respond appropriately to parental alarm calls, these results demonstrate that nest predation by multiple predators can drive evolution of complex parent–offspring communication in altricial species.
Low frequency calls of cassowaries -- Although some birds can detect wavelengths in the infrasound range,there has been litle evidence that birds produce very low frequencies. Mack and Jones (2003) made 9 recordings of a captive Dwarf Cassowary (Casuarius benneti) and one recording of a wild Southern Cassowary (C. casuarius) in Papua New Guinea. Both species produced sounds near the floor of the human hearing range in their pulsed booming notes: down to 32 Hz for C. casuarius and 23 Hz in C. benneti.
Natural selection should favor the evolution of vocalizations that reach their targets with minimal degradation, and low frequencies propagate over long distances with minimal attenuation caused by vegetation. New Guinea forests ofen have a fairly thick understory of wet leafy vegetation that could quickly attenuate higher frequencies. Thus, the very low frequency calls of cassowaries probably ideal for communication among widely dispersed, solitary cassowaries in dense rainforest. How cassowaries produce such low vocalizations is currently unknown.
All three cassowary species have keratinous casques rising from the upper mandible over the top of the skull up to 17 cm in height. Hypotheses concerning the function of the casque include: (1) a secondary sexual character, (2) a weapon in dominance disputes, (3) a tool for scraping the leaf-litter, or (4) a crash helmet for birds as they bash through the undergrowth. The later three seem unlikely based on field observations. Future research should include the possibility that the casque might play some role in sound reception or acoustic communication.
.
Photo by D. DeMello, Wildlife Conservation Society
The functions of bird song may vary among species. Some known & hypothesized functions include:
Identification:
Songs have characteristics that permit other birds to identify the species, sex (if both males and females sing), and individual identity of a singer.
Characteristics important in permitting specific, sexual, & individual recognition vary among species but may include (Becker 1982):
song duration
interval between song elements (also called notes or syllables)
frequency
syntax - the order of elements within a song
structure of elements, e.g., duration and frequency
Mate attraction
Responses of female Lincoln's Sparrows vary with male song quality -- Sexually reproducing organisms should mate with the highest quality individuals that they can. When female songbirds choosea mate, they are thought to use several aspects of male song that reflect his quality. Under resource-limited environmental conditions, male Lincoln’s Sparrows (Melospiza lincolnii)vary among one another in several aspects of song quality, includingsong length, song complexity, and trill performance. In a 2-prongedapproach, Caro et al. (2010) tested whether variation in song quality of maleLincoln’s Sparrows influences the behavior of femalesthat are in a reproductive-like state. Over two trials, females were exposed to songs from the high and low ends of the distributionof naturally occurring song quality variation and were found to exhibit a higherlevel of behavioral activity when exposed to high-qualitysongs, especially females that had first been exposed to low-qualitysongs. Female phonotaxis toward antiphonallyplayed songs was also examined with experimentally elevated and reduced trillperformance, and females were found to move preferentially towardthe songs with elevated trill performance. These results demonstratethat the behavior of female Lincoln’s Sparrows is modulatedby the quality of male songs to which they are exposed and thattrill performance plays a significant role in this behavioralmodulation. Furthermore, as the order of song quality presentationmatters, it appears that recent song experience also influencesfemale behavior.
Pairing success was measured by relative pairing date. Birds pairing relatively late are those that demonstrate longer intervals (in days) between arrival and the start of egg laying than birds realizing quick pairing. Data are residuals from a multiple regression, in which the date of arrival was included to control for date effects.
Birds reveal their personality when singing -- Individual differences in social behavior may have consequences for mate choice and sexual signalling, because partners should develop preferences for personalities that maximize reproductive output. Here we propose that behavioural traits involved in sexual advertisement may serve as good indicators of personality, which is fundamental for sexual selection to operate on temperament. Bird song has a prominent and well-established role in sexual selection, and it displays considerable variation among individuals with a potentially strong personality component.
In a field study of free-living male Collared Flycatchers (Ficedula albicollis), Garamszegi et al. (2008) characterized personality based on the exploration of an altered breeding environment, and based on the risk taken when a potential predator was approaching during a simulated territorial interaction. They found that explorative and risk-taker individuals consistently sang at lower song posts than shy individuals in the presence of a human observer. Moreover, males from lower posts established pair-bonds relatively faster than males from higher posts.
These results may demonstrate that risk taking during singing correlates with risk taking during aggression and with exploration, thus personality may be manifested in different contexts involving sexual advertisement. These findings are in accordance with the hypothesis that the male's balance between investment in reproduction and risk taking is reflected in sexual displays, and it may be important information for choosy females that seek partners with personality traits enhancing breeding success.
Side, front and back thermal images of a singing canary. The images are colour coded to show temperature
increasing from 15°C (dark blue) through green (22°C), yellow (25°C) and red (30°C) to 32°C (white). The black lines
delineate the head, body, leg and tail sections of the bird (From: Ward and Slater 2005).
Energetic cost of singing - Sexually selected displays, such as male passerine bird song, are predicted to be costly. However, most measurements calculated the rate of oxygen consumption during singing using respirometry have shown that bird song has a low energetic cost. Because birds are reluctant to sing when enclosed inside a respirometry chamber, the energetic cost of singing could differ from that under more normal circumstances. Ward and Slater (2005) used heat transfer modelling, based on thermal images, to estimate the energetic cost of singing by Canaries (Serinus canaria) that were not enclosed in respirometry chambers. Metabolic rate calculated from heat transfer modelling was 14% greater than during standing, suggesting that song production appears to be metabolically cheap for passerine birds and the metabolic cost small enough that it is unlikely represents an important fitness cost under circumstances favorable for energy balance. However, cost will increase as the temperature decreases.
Territory establishment and defense
Motivation and Fitness - Birds may provide information to conspecifics by way of variation in (Becker 1982):
singing rates
may increase during aggressive encounters
may be higher in higher quality males
song duration - may increase or decrease (depending on the species) during conflict situations
song amplitude (or volume) may decrease during aggressive encounters
song frequency - may increase during conflict situations (e.g., Indigo Buntings; Thompson 1972)
song complexity - songs may consist of more or fewer elements during conflict situations (e.g., male Blue Grosbeak utter songs with more syllables during aggressive encounters with other males)
Distraction of potential predators (e.g., Common Yellowthroat flight song)
Coordination of activities
Stimulate females
Attract females for extra-pair copulations
Mate guarding
Song complexity and the avian immune system -- There are three hypotheses to explain how the evolution of parasite virulence could be linked to the evolution of secondary sexual traits, such as bird song. First, female preference for healthy males in heavily parasitized species may result in extravagant trait expression. Second, a reverse causal mechanism may act, if sexual selection affects the coevolutionary dynamics of host-parasite interactions by selecting for increased virulence. Third, the immuno- suppressive effects of ornamentation by testosterone or limited resources may lead to increased susceptibility to parasites in species with elaborate songs. Assuming a coevolutionary relationship between parasite virulence and host investment in immune defense, Garamszegi et al. (2003) used measures of immune function and song complexity to test these hypotheses in passerine birds. Under the first two hypotheses, they predicted avian song complexity to be positively related to immune defense among species, whereas this relationship was expected to be negative if immuno-suppression was at work. They found that adult T-cell mediated immune response and the relative size of the bursa of Fabricius were both positively correlated with song complexity, even when potentially confounding variables were held constant. These results are consistent with the hypotheses that predict a positive relationship between song complexity and immune function, thus indicating a role for parasites in sexual selection.
Regression of short-term song complexity (number of
unique syllables within songs/song length) on T-cell
mediated immune response, after removing allometric
effects by using residuals after controlling for body
mass. Datapoints are phylogenetically independent
linear contrasts (N = 38). The line and equation are from
linear regression forced through the origin.
Song differences of Great Tits at urban and forest sites. (A) Every urban site had a higher minimum frequency than its forest companion site. (B) The minimum frequency (+ SE) varies with note number, and the habitat-dependent spectral divergence remained distinct irrespective of number of notes. Squares = songs from cities; triangles = songs from forest. (C) Minimum frequency correlated between sites of a pair without a strong, larger-scale geographic pattern of isolation by distance. City-forest pairs are labeled by city name. The line of equal values for city-forest pairs in is the top-left corner. (D) Every urban site had a shorter note length than its forest companion site. (E) The duration of the first note varied with note number, but the habitat-dependent temporal divergence remained distinct, especially for the note numbers with substantial sample sizes. Squares = songs from cities; triangles = songs from forest. (F) Duration of the first note did not correlate between sites of a pair, nor was there a larger-scale geographic structure. City-forest pairs are labeled by city name. The line of equal values for city-forest pairs in is the bottom-right corner.
Cities change the songs of birds -- Worldwide urbanization and the ongoing rise of urban noise levels form a major threat to living conditions in and around cities. Urban environments typically homogenize animal communities, and this results, for example, in the same few bird species' being found everywhere. Insight into the behavioral strategies of the urban survivors may explain the sensitivity of other species to urban selection pressures. Slabbekoorn and den Boer-Visser (2006) showed that songs that are important to mate attraction and territory defense have significantly diverged in Great Tits (Parus major), a successful urban species. Urban songs were shorter and sung faster than songs in forests, and were often atypical song types. The authors also found consistently higher minimum frequencies in ten out of ten city-forest comparisons from London to Prague and from Amsterdam to Paris. Anthropogenic noise is most likely a dominant factor driving these dramatic changes. These data provide the most consistent evidence supporting the acoustic-adaptation hypothesis since it was postulated in the early seventies. At the same time, they reveal a behavioral plasticity that may be key to urban success and the lack of which may explain detrimental effects on bird communities that live in noisy urbanized areas or along highways.
In some species, females also sing. This is particularly true in the tropics (see 'duetting' below). Singing by females may be important in:
territory defense (particularly in keeping other females out of a territory)
pair-bonding
mate guarding
reproductive synchronization
When do female birds sing? Hypotheses from experimental studies (Langmore 1998).
(a) The song of a female Superb Fairy-wren. Females use songs to defend their territories against both
males and females. (b) The song of a female Alpine Accentor. Female Alpine Accentors sing to attract males, and
the complexity of their songs increases with age. This song was recorded from a two-year-old female (Langmore 1998).
Seminar - Divas in the treetops: when and why do female birds sing? by Lauryn Benedict
Want to know more about singing by female songbirds? Check femalebirdsong.org
Song function scores with levels of support. Blue bars represent studies where authors confirmed female song function for their study species. Red bars represent studies where authors refuted female song function for their study species. Each support level is stacked, where bars with the darkest color represent studies that confirmed or refuted a given function category based on fitness testing, those of medium color represent behavioural testing and those with the lightest color represent hypothesized conclusions about each song function category (From: Barros et al. 2024).
Singing by female Northern Cardinals -- Yamaguchi (2001) found that female Northern Cardinals learn to sing three times faster than males - the most dramatic example of learning disparities between male & female animals found to date. She collected nestling cardinals & raised them in sound chambers with microphones and speakers that play back the songs of adult cardinals. It takes about a year for a cardinal to learn to sing, and young songbirds learn by imitating adults. During the early sensitive phase, young don’t sing, but listen to singing adults to memorize their songs. Then the practicing begins. “Their initial attempts are pretty miserable,” she says, “but they practice & practice until it matches the memory that was formed earlier during the sensitive phase.”
Yamaguchi (1998) also analyzed songs and found that females sing with more overtones, creating a slightly nasal sound. Young males also go through a nasal, warbly phase as their testosterone kicks in, but it’s as though the females continue to sing with an adolescent male’s voice. More important, Yamaguchi (2001) discovered that female cardinals memorize adult songs three times faster than males. While both sexes ultimately learned the same number of song types, the females’ sensitive phase was only a third as long as the males’. The different learning rates may reflect an evolutionary adaptation. Like other songbirds, juvenile cardinals disperse from their parents’ territory about 45 days after hatching to establish their own turf before their first breeding season. Away from their natal nest, the young cardinals are suddenly immersed in the new song dialects of other adult cardinals. It appears that females lose the ability to learn new dialects when they disperse, while males are able to learn them and “fit in” with their new neighbors.
“ It might be that males retain the ability to learn songs longer than females so that they can have a better chance of establishing territory in a new area,” Yamaguchi says. “For males, song-matching and fitting into the crowd in a new place are really important, while they’re not for females.” It’s not clear why female cardinals have a shorter window of vocal learning, she says, but then again, “we don’t really know why females sing at all, or how they use their songs.” One hypothesis, she says, is that females sing as a species identification tool, a greeting card to male cardinals that says, “I’m an eligible mate; come court me.” Other researchers have proposed that female cardinals sing to shoo away brightly colored mates from the nest, warning the males not to attract attention to the vulnerable chicks. “I know that female cardinals also use songs in aggressive behavior,” Yamaguchi says. “I’ve seen females battling each other in the field, and they’re singing the whole time as they bang into each other.”
Singing male Northern Cardinal
Early bird: the European Robin starts singing over an hour before some of its neighbors (Photo by S. Crawford)
Early birds have big eyes -- Birds with large eyes begin singing earlier in the morning than their small-eyed neighbors because they can see better in low light. So say British researchers explaining why, on a spring day in Welsh woodland, robins and redstarts pipe up a good hour and a half before chaffinches and blue tits. The keen-sighted early birds probably do get the worms. Big eyes may help the hunt for breakfast, speculates Robert Thomas from the University of Bristol, who led the study (Thomas et al. 2002). Birdsong is an acoustic advertisement. Birds use it to attract mates and defend territory. But singing also alerts predators. So a bird does not sing before it can see well enough to spot approaching danger, speculates Thomas. Yet a bird also does not want to waste time crooning when the sun is high enough to search for food. The earlier it can see to sing, the earlier it can see to start foraging. The link between the dawn chorus and eye size was first mooted in the 1960s, but this is the first time that it has been tested. Previous data was very confusing, says Graham Martin, who works on birds' senses at the University of Birmingham, UK. Thomas's team measured light levels and the times at which 57 different bird species began singing at various sites across Britain and Europe. When they measured the diameter of each bird's exposed eye surface, they realized that big-eyed birds sing earlier than those of the same body size but with smaller eyes. They also found that small birds sing earlier than large birds with the same-sized eyes as them. One explanation for this may be hunger. Small birds cannot store as much food overnight and they use up energy more quickly than large birds. "Only large birds can afford to take it easy," says Thomas Szekely, another member of the research team. "Small birds may sing earlier because they have to get up earlier to search for food." -- Natasha McDowell, Nature Science Update
Song Variation
Locally distinct versions of song (or song 'dialects') have been described in several species. Hypotheses to explain dialects include:
Historical hypothesis - Because vocal dialects are the result of song learning, geographical variation in song may simply reflect recent history. New 'dialects' develop when birds (who sing particular songs or song types) colonize new areas (Gill 1995).
Racial specialization, or ecological, hypothesis - Song dialects reflect the genetics of local populations. Females may make mate choice decisions based on a male's dialects and young males may establish territories in particular areas based on the dialects of resident male. If so, dialect boundaries may represent 'genetic' boundaries, with birds in a particular 'dialect area' genetically 'adapted' to that particular area. This may be the case in certain populations of White-crowned Sparrows in the western United States.
Social adaptation hypothesis - Dialects develop when young males dispersing into an area learn the songs of established males. For example, in Indigo Buntings, young males that sing like their neighbors have greater reproductive success (Payne 1982).
Song Repertoires
In many species of songbirds, including Northern Cardinals, Carolina Wrens, and many others, males possess repertoires of song types. These repertoires typically consist of perhaps 6 - 12 song types, but may range anywhere from 2 to an apparently unlimited number (as in, for example, Northern Mockingbirds). Interestingly, however, males in other species, such as Common Yellowthroats, have just one song. Repertoires of song types may have a variety of functions:
Each song type has a different meaning
Advertise quality (more song types = better quality)
Permit more effective communication with conspecific males (i.e., matching the song types of neighbors)
Prevent 'exhaustion' of nerves & muscles
Prevent habituation
Relationships between a male song sparrow's song repertoire size and mean kinship (km) with the female population. Males with larger song repertoires were less closely related to the female population, including all females in the set of potential mates (open symbols, dashed line) and excluding close female relatives of each focal male (filled symbols, solid line). Regression lines are shown for clarity.
Secondary sexual ornamentation and non-additive genetic benefits of female mate choice -- Ornamental secondary sexual traits are hypothesized to evolve in response to directional mating preferences for more ornamented mates. Such mating preferences may themselves evolve partly because ornamentation indicates an individual's additive genetic quality (good genes). While mate choice can also confer non-additive genetic benefits (compatible genes), the identity of the most 'compatible' mate is assumed to depend on the choosy individual's own genotype. It is therefore unclear how choice for non-additive genetic benefits could contribute to directional mating preferences and consequently the evolution of ornamentation. In free-living Song Sparrows (Melospiza melodia), Reid (2007) found that individual males varied in their kinship with the female population. Furthermore, a male's song repertoire size, a secondary sexual trait, was negatively correlated with kinship such that males with larger repertoires were less closely related to the female population. The smaller repertoires of more closely-related males probably represents direct inbreeding depression in the expression of this secondary sexual trait. After excluding close relatives as potential mates, individual females were on average less closely related to males with larger repertoires. Therefore, female Song Sparrows expressing directional preferences for males with larger repertoires would on average acquire relatively unrelated mates and produce relatively outbred offspring. Such non-additive genetic fitness benefits of directional mating preferences, which may reflect genetic dominance variance expressed in structured populations, should be incorporated into genetic models of sexual selection.
Song sharing (Beecher and Brenowitz 2005) -- Song sharing is common in songbirds and is found in a variety of social contexts, not only in territorial neighbors (the most commonly studied context), but also in lekking species and communal breeders. Song sharing in the neighbor context is best understood in the context of the Dear Enemy hypothesis. According to this hypothesis, long-term neighbors are preferred to newcomers because newcomers are inherently expansionist, whereas old neighbors generally respect territory boundaries once they have been mutually established. Neither preferring nor cooperating with familiar neighbors requires shared songs, but shared songs are a reliable signal (a ‘badge’) of familiarity or locality because they must be learned in the local neighborhood. Consistent with this hypothesis, Wilson and Vehrencamp (2001) found that neighboring Song Sparrows sharing fewer songs were more aggressive with one another than were neighbors sharing more songs.
A corollary of the Dear Enemy hypothesis for territorial songbirds is that established neighbors should use their songs in place of time- and energy-costly physical interactions to minimize unnecessary territorial conflicts. Playback studies of Song Sparrows and Banded Wrens have supported this prediction. Even when neighbors do not share any song types (with respect to the investigators' criterion), they might still be able to song match using songs in their repertoires that they perceive as being most similar.
Song learning
Avian vocalizations can be inherited, learned, or invented. In some species, like Brown-headed Cowbirds, males develop normal songs even when raised in acoustic isolation. However, for most species of songbirds, at least some aspects of their singing behavior is learned. That learning, though, is guided by inherited (innate) mechanisms.
Stages of song development (birds with fixed repertoires):
1 - Critical learning period
information stored for later use
usually less than a year in duration, & may be much shorter than that (e.g., 2 months for Marsh Wrens)
often extends into the 1st breeding season (giving young males a chance to learn from experienced, neighboring males)
2 - Silent period
song components that have been memorized during learning period are stored in the brain
up to 8 months in duration
3 - Subsong period
period of practice without communication
birds utter low volume, unstructured notes initially and, in a few weeks, sounds become more like the species-specific song
first attempts to sing (plastic song); birds continue to alter song structure
4 - Song crystallization
plastic song becomes increasingly less 'plastic' and, finally, the 'adult (or final) version' of song(s) is(are) sung
in crystalized song, birds may only use a fraction of the elements they first learned and practiced
Stages and duration of song development in White-crowned Sparrows
Timeline for Zebra Finch song learning. Young birds first memorize the song of an adult 'tutor' during a
critical period for 'sensory' learning of song. Later, during a 'sensorimotor' learning period, they begin to sing and
gradually match their initially immature vocalizations to the memorized song using auditory feedback. After this learning
process, adult song normally remains unchanging, or 'crystallized'. For Zebra Finches, the periods of
sensory and sensorimotor learning overlap. Many other species, such as the White-crowned Sparrow, do not crystallize
song until close to one year of age, and have a much clearer separation between the sensory and sensorimotor
learning periods. Still other species, such as the canary, seem to reiterate the processes of sensory and sensorimotor
learning each season, perhaps under the regulation of seasonal variability in hormone levels (Brainard and Doupe 2000).
Photomicrographs showing HVC of control (A) and parasitized (B) male Canaries and Robust nucleus of theArchopallium (RA) of control (C)
and parasitized (D) birds. HVC was smaller in the parasitized birds (B) compared to the control(A) group, whereas RA showed no reduction in size (C vs. D). Scale bar is 100 mm.
Parasites affect song complexity and neural development in a songbird -- There is now considerable evidence that female choice drives the evolution of song complexity in many songbird species. However, the underlying basis for such choice remains controversial. The developmental stress hypothesis suggests that early developmental conditions can mediate adult song complexity by perturbing investment in the underlying brain nuclei during their initial growth. Spencer et al. (2005) found that adult male Canaries (Serinus canaria), infected with malaria (Plasmodium relictum) as juveniles, develop simpler songs as adults compared to uninfected individuals, and exhibited reduced development of the high vocal center (HVC) song nucleus in the brain. Their results show how developmental stress not only affects the expression of a sexually selected male trait, but also the structure of the underlying song control pathway in the brain, providing a direct link between brain and behavior. This novel experimental evidence tests both proximate and ultimate reasons for the evolution of complex songs and supports the Hamilton–Zuk hypothesis of parasite-mediated sexual selection. Together, these results propose how developmental costs may help to explain the evolution of honest advertising in the complex songs of birds.
Singing in their sleep - Researchers have shown that young birds sleeping at night may be reviewing the songs they've learned during the day (Dave and Margoliash 2000). Normally, the brain is desensitized to outside stimuli during sleep, partly because of changing concentrations of a norepinephrine. But the robustus archistratalis (RA), a region of the Zebra Finch brain involved in singing, exhibits an increase in neurological activity during sleep. It is generally believed that the activity of the sleeping brain helps to consolidate what was learned during the day, but how this occurs has never been directly shown. Dave and Margoliash (2000) recorded electrical impulses from single neurons in the RA of anesthetized, asleep and awake birds as they listened to recordings of their own songs played back on a computer. Without fail, birds that were asleep or anesthetized exhibited reduced regular oscillations but showed occasional bursts of strong activity in their RA neural impulse patterns. When the birds occasionally woke up during the night, the bursting patterns quickly disappeared and were replaced by the steady oscillating pattern seen during the day. "This is surprising because the same neurons that show no response during the day have these strong responses to the bird's own song when they are asleep. It's possible that songs learned during the day affect the bursting patterns of the RA at night, serving to solidify the newly learned songs in the bird's mind," says Margoliash.
a,Left and bottom: schematic of the Zebra Finch song system, experimental protocol and timeline of the experiments. Inset,Nissl-stained image of HVC in parasagittal section, showing its location on the floor of the lateral telencephalic ventricle, ~100μm below the pial surface. RA,robust nucleus of the arcopallium; areaX,striatal component of the song system; scale bar,200μm. Right: in vivo two-photon image of GFP-labelled spinous HVC neurons amid retrogradely labelled RA-projecting (HVCRA,red) and area-X-projecting (HVCX,blue) neurons. Scale bar,20μm. b,Repeated in vivo imaging of dendritic branches from an HVC neuron of a 130-d zebra finch over several days. The rightmost three images (scale bar,10μm) show the boxed region in the leftmost image (scale bar,20μm) at the times indicated. c,Views of the dendritic segment shown boxed in the 0-h image inb, imaged 2h apart. d,View of another dendritic segment of an HVC neuron, showing the gain and loss of dendritic spines across a 2-h imaging interval. In c and d,arrowheads point to stable (blue), lost (yellow) and gained (green) spines. Scale bars,2μm.
Synaptic enhancement during song learning -- Behavioral learning depends on the brain’s capacity to respond to instructive experience and is often enhanced during a juvenile sensitive period. How instructive experience acts on the juvenile brain to trigger behavioral learning remains unknown. In vitro studies show that forms of synaptic strengthening thought to underlie learning are accompanied by an increase in the stability, number and size of dendritic spines, which are the major sites of excitatory synaptic transmission in the vertebrate brain. In vivo imaging studies in sensory cortical regions reveal that these structural features can be affected by disrupting sensory experience and that spine turnover increases during sensitive periods for sensory map formation. These observations support two hypotheses: first, the increased capacity for behavioural learning during a sensitive period is associated with enhanced spine dynamics on sensorimotor neurons important for the learned behavior; second, instructive experience rapidly stabilizes and strengthens these dynamic spines. Roberts et al. (2010) report a test of these hypotheses using two-photon in vivo imaging to measure spine dynamics in Zebra Finches, which learn to sing by imitating a tutor song during a juvenile sensitive period. Spine dynamics were measured in the forebrain nucleus HVC, the proximal site where auditory information merges with an explicit song motor representation, immediately before and after juvenile finches first experienced tutor song. Higher levels of spine turnover before tutoring correlated with a greater capacity for subsequent song imitation. In juveniles with high levels of spine turnover, hearing a tutor song led to the rapid (~24-h) stabilization, accumulation and enlargement of dendritic spines in HVC. Moreover, in vivo intracellular recordings made immediately before and after the first day of tutoring revealed robust enhancement of synaptic activity in HVC. These findings suggest that behavioral learning results when instructive experience is able to rapidly stabilize and strengthen synapses on sensorimotor neurons important for the control of the learned behavior.
What is learned?
1 - Syllables?
innate in Galliforms, Columbiformes, and suboscines
learned in most oscines studied, but some species can derive syllables from call notes (e.g., begging calls)
2- Duration?
heritability of duration is evident in song of domestic non-oscines that have been selected for different song lengths (e.g., crowing duration in domestic chickens and cooing duration of domestic pigeons)
even among songbirds that learn songs, duration appears to be constrained, e.g., song durations of isolate-reared and tutored White-crowned Sparrows do not differ
3 - Rhythm?
duration of notes and intervals between them that are characteristic of species
fixed in non-oscines and apparently in many species that learn songs, e.g., Song Sparrow's songs begin with a phrase containing notes that accelerate in tempo and the same rhythm appears in isolate-reared and tutored birds
4 - Frequency?
dispositions to produce sounds of certain frequencies may be under direct genetic control, but mean frequencies and ranges are often correlated with body size, e.g., vocal frequency is inversely proportional to mass in adult Northern Bobwhites
related to characteristics (e.g., thickness) of syringeal membranes
Male Ability To Learn Song Affects Female Mating Response -- Researchers have found the first evidence that female birds use song-learning ability as an indicator of male quality. Nowicki et al. (2002) tested the response of female Song Sparrows to songs of captive-raised males and found that females preferred songs that came closest to wild-type songs they heard when young and presumably learned as models. Nowicki & his colleagues have long theorized that female songbirds pay attention to male song as an indicator of fitness. "We've developed experimental evidence that there is a link between early stress, male brain development and song-learning," he said. "But until now, experimental and field observations showing that females were interested in song only contrasted the presence or absence of song, or relatively gross features of song, like the size of the repertoire. This is the first study to demonstrate that females care about song-learning quality," he said. To test the effects of fine differences in song quality on female response, the researchers trained captive-reared male song sparrows to sing by exposing them to the recorded songs of wild birds. To induce variation in stress among the birds, some were placed on a restricted diet during development. Using spectrographic analysis, the researchers rated the captive-reared birds on two measures of song quality: (1) how much of the wild-bird song they copied versus how much they invented, a practice common among Song Sparrows. Those birds who did invent more song elements also tended not to copy well those elements they did copy, and (2) how close the males had come to actually matching just the wild-bird song elements they were attempting to copy. To determine the effects of song quality, the researchers exposed wild-caught adult females to the captive-reared males' songs & measured female response to the songs, including shivering of the wings, lifting of the tail and a characteristic call. As a control, the wild-caught females were exposed to what the scientists had judged as well-learned male songs, as well as the digitally recorded wild songs. The female birds responded equally to both. However, when exposed to the captive-reared males' songs, females responded more strongly to male songs that had been better learned by both of the scientists' measures. "The females showed a strong preference for songs that had been copied well, as opposed to songs that had been copied poorly," said Nowicki. "And by our measures, the males got points taken off for originality. That seems to make sense because we would argue that males that deviate from original song haven't learned the song as well."
"We know sexual selection is a very powerful evolutionary force that has led to phenomena such as the evolution of extravagant displays and the evolution of size differences between sexes", said Nowicki, "I believe that this work demonstrates that sexual selection might not be acting directly on the obvious trait that is expressed, but on the mechanisms that underlie the expression of that trait. In the case of bird song, a male's song reflects the birds' developmental history, and song expression is only the trait that the female can gain access to for information about -- in this case -- brain mechanisms." While the current studies show clearly that females prefer well-learned songs, among the next research steps, said Nowicki, will be to determine how females learn to judge song quality. "There is only very thin evidence that females learn song, so it's a major scientific question whether females are learning something about the population that they're living in, and using that as a way of assessing males," he said. Such female studies also will reveal whether the female's ability to distinguish good songs from bad reflects the birds' fitness and influences evolution, said Nowicki. (Source: Science Daily)
Why have songbirds evolved the ability to learn their songs?
vocal learning & associated neural control system arose only once in the Passeriformes (in ancestor of modern songbirds but not in the ancestral suboscine)
selective advantage?
larger song repertoires
learned song dialects
Wide repertoire wins mates -- Female Great Reed Warblers choose males who sing the widest repertoire of songs because it shows they were well brought up. Songs are learned at a vulnerable stage of fledgling development, so Nowicki et al. (2000) predicted that adult males with the most tunes were well fed as youngsters -- turning them into eligible bachelors. Although biologists know that male birdsong attracts females, the exact message it conveys is a mystery. "We just don't know how the male bird song can mean anything to the female," Nowicki explains. He now claims to have the first experimental data that can answer this question. "A male's songs may be an honest indicator of how well he developed in the face of nutritional or other stresses experienced early in life," he suggests. Hungry chicks, in other words, have more on their mind than choir practice. And undernourished fledglings generally make poorer fathers when they grow up. To test the idea, Nowicki et al. (2000) recorded year-old Great Reed Warblers (Acrocephalus arundinaceus) in Sweden; they also weighed the birds and measured the lengths of particular feathers -- the longest of which are usually found on the strongest, fittest birds. Warblers with the most warbles also had the longest feathers -- supporting Nowicki's idea. The researchers also found evidence of a link between wide song repertoire and body mass. So, what do female warblers think when they encounter males with limited songs? That they are being chatted up by real bird-brains, Nowicki says. Limited song learning indicates poorer brain development in general. "By assessing the output of song learning, the female may gain accurate information about critical cognitive abilities such as spatial navigation and memory," he says. These are important for skills that every good warbler father needs: the ability to defend territory, for example, avoid predators and find food. This argument provides "a convincing solution to the problem," says Nigel Mann of the bird and mammal sound communication group at St. Andrews University, UK. "Other studies have shown that ability in tits to remember where food stores are located correlates with the size of certain brain nuclei. An interesting further development of the idea, " Mann continues, "would be to look more directly for a connection between such aspects of foraging, spatial memory and repertoire size." -- David Adam, Nature Science Update
Male Great Reed Warbler
(A, B) Problem-solving ability grouped by vocal learning category (closed-ended [including non-vocal learning species], open-ended vocal learning species, and mimics vs. non-mimics). (C) Relationship between vocalization repertoire size (songs and calls combined) and problem-solving ability for 214 individuals of 23 species of birds (21 songbirds and two non-songbirds). Problem-solving ability was determined using four different obstacle-removal tasks, i.e., obstacles had to be moved to gain access to a food reward. The average number of trials required to solve the four tasks was used as the measure of problem-solving performance. Different symbols represent the different species of birds.
Complex vocal learning has been assumed to be associated with more-advanced cognitive abilities. However, tests of this hypothesis have been inconclusive and have not been conducted across species. Audet et al. (2023) measured an array of cognitive skills, including problem-solving, across 214 individuals of 23 bird species, including 19 wild-caught songbird species, two domesticated songbird species, and two wild-caught vocal non-learning species.They found that the greater the vocal learning abilities of a species, the better their problem-solving skills and the relatively larger their brains. These conclusions held when controlling for non-cognitive variables and phylogeny. These results support a hypothesis of shared genetic and cognitive mechanisms between vocal learning, problem-solving, and bigger brains in songbirds.
Vocal mimicry, in which birds copy and imitate sounds produced by other birds (or just other sounds in general), occurs in about 15-20% of passerines. Among the best examples of 'mimics' are birds in the family Mimidae (Northern Mockingbird, Brown Thrasher, and Gray Catbird). The possible functions of mimicry include:
Intraspecific functions - imitation is a way to increase song complexity and more complex singing may help males:
attract mates
better defend territories
stimulate mates to bring them into reproductive condition
Interspecific functions:
interspecific territoriality - another species is unlikely to be deceived by mimicked song but may learn to associate mimicked song with an aggressive 'mimic' (e.g., a Northern Mockingbird aggressively defending a food source)
overlapping bouts of vocal or non-vocal sounds given by members of a mated pair
typically associated with tropical birds that exhibit year-round territoriality & prolonged monogamous pair bonds
Possible functions of duetting:
Territorial
Pair-bonding
Mate guarding
Neural mechanisms of coordination in duetting wrens by Melissa Coleman
Complex song duet of the Plain Wren -- Mann et al. (2003) studied the duet of the Caribbean-slope subspecies of the Plain Wren (Thryothorus modestus zeledoni) in Costa Rica. It is one of the most complex duets to have been described. The duet proper consists of rapid, highly coordinated alternation of “A-phrases” from the female and “B-phrases” from the male. While the female initiates this section with her A-phrase, this cyclical part of the duet is almost invariably preceded by an introductory “I-phrase” from the male, so that it is the male that initiates the performance. Each male has a repertoire of I- and B-phrases, and each female has a repertoire of A-phrases. These are specifically associated with each other to form a repertoire of duet types. Mann et al. (2003) hypothesize that the pattern of song organization in this species facilitates more coordinated and precise duetting. The presence of the three components means that a full duet requires the cooperation of both members of the pair. Some investigators believe that duets benefit both singers, and play a role in territorial defense, pair bonding, or maintenance of contact. Other authors believe a conflict of interests may be involved, e.g., a female whose mate is attempting to attract a second female might benefit by labelling him as already being mated. While a purely descriptive study such as this one cannot resolve such issues, highly complex and tightly coordinated duets such as those of the Plain Wren seem more likely to stem from cooperation than conflict.
Standard duet format of the Plain Wren. A typical duet, initiated by a male introductory (I-) phrase and followed by an antiphonal cycle of female (A-) and male (B-) phrases. Recorded during October 2001 at La Suerte, Costa Rica.
Arnold, A. P. 1980. Sexual differences in the brain. American Scientist 68: 165–173.
Arnold, A. P., F. Nottebohm, and D.W. Pfaff. 1976. Hormone concentrating cells in vocal control and other brain regions of the Zebra Finch (Poephila guttata). J. Comp. Neurol. 165: 487–512.
Audet et al. 2023. Songbird species that display more-complex vocal learning are better problem-solvers and have larger brains. Science 381: 1170-1175.
Barros et al. 2024. A review of the literature on female birdsong function. Animal Behaviour 216: 23-35.
Becker, P.H. 1982. The coding of species-specific characteristics in bird sounds. Pp. 213-252 in Acoustic Communication in Birds, vol. 1 (D.E. Kroodsma and E.H. Miller, eds.). Academic Press, New York, NY.
Beecher, M. D. and Eliot A. Brenowitz. 2005. Functional aspects of song learning in songbirds. Trends in Ecology and Evolution 20:143-149.
Bentley, G.E., T. Van't Hof, & G. F. Ball. 1999. Seasonal neuroplasticity in the songbird telecephalon: A novel role for melatonin. Proceedings of the National Academy of Sciences USA 96: 4674-4679.
Brainard, M.S. and A.J. Doupe. 2000. Auditory feedback in learning and maintenance of vocal behavior. Nature Reviews Neuroscience 1:31-40.
Brenowitz, E. A. and K. Lent. 2002. Act locally and think globally: Intracerebral testosterone implants induce seasonal-like growth of adult avian song control circuits. Proc. Natl. Acad. Sci. 99:12421-12426.
Caro, S. P., K. B. Sewall, K. G. Salvante, and K. W. Sockman. 2010. Female Lincoln's Sparrows modulate their behavior in response to variation in male song quality. Behavioral Ecology 21: 562569.
Dave, A.S. and D. Margoliash. 2000. Song replay during sleep and computational rules for sensorimotor vocal learning. Science 290: 812-816.
Duffy, D.L. and G. F. Ball. 2002. Song predicts immunocompetence in male European starlings (Sturnus vulgaris). Proc. Roy. Soc. B 269:847-852.
Elemans, C. P. S., I. L. Y. Spierts, U. K. Muller, J. L. van Leeuwen, and F. Goller. 2004. Superfast muscles control dove's trill. Nature 431: 146
Garamszegi, L. Z., M. Eens, and J. Török. 2008. Birds Reveal their Personality when Singing. PLoS ONE 3(7): 0002647.
Garamszegi, L. Z., A. P. Møller, and J. Erritzøe. 2003. The evolution of immune defense and song complexity in birds. Evolution 57: 905-912.
Gill, F.B. 1995. Ornithology, 2nd ed. W.H. Freeman and Co., New York, NY.
Goller, F. and O. N. Larsen. 1997a. A new mechanism of sound generation in songbirds. Proc. Natl. Acad. Sci. USA 94: 14787-14791.
Goller, F. and O. N. Larsen. 1997b. In situ biomechanics of the syrinx and sound generation in pigeons. Journal of Experimental Biology 200:2165-2176.
Goller, F. and O.N. Larsen. 1999. Role of syringeal vibrations in bird vocalizations. Proc. Roy. Soc. London B 266:1609.
Griesser, M. 2008. Referential calls signal predator behavior in a group-living bird species. Current Biology 18: 69-73.
Konishi, M. and E. Akutagawa. 1985. Neuronal growth, atrophy, and death in a sexually dimorphic song nucleus in the zebra finch brain. Nature 315: 145–147.
Laje, R., T. Gardner and G. B. Mindlin. 2002. Neuromuscular control of vocalization in bird song: a model. Phys. Rev. E 65: 051921.
Langmore, N. E. 1998. Functions of duet and solo songs of female birds. Trends in Ecology and Evolution 13:136-140.
Mack, A. L. and J. Jones. 2003. Low-frequency vocalizations by Cassowaries (Casuarius spp.). Auk 120: 1062-1068.
Mann, N. I., L. Marshall-Ball, and P.J. B. Slater. 2003. The complex song duet of the Plain Wren. Condor 105: 672-682.
Neudorf, D. L. and S. G. Sealy. 2002. Distress Calls of Birds in a Neotropical Cloud Fores. Biotropica 34: 118-126.
Nottebohm, F. 1980. Testosterone triggers growth of brain vocal control nuclei in adult female canaries. Brain Res. 189: 429–436.
Nottebohm, F. 1981. A brain for all seasons: cyclical anatomical changes in song control nuclei of the canary brain. Science 214: 1368–1370.
Nowicki, S., D. Hasselquist, S. Bensch & S. Peters. 2000. Nestling growth and song repertoire size in great reed warblers: evidence for song learning as an indicator mechanism in mate choice. Proceedings of the Royal Society B 267:2419 - 2424.
Nowicki, S., W.A. Searcy, and S. Peters. 2002. Quality of song learning affects female response to male bird song. Proc. Roy. Soc. B 269:1949-1954.
Payne, R.B. 1982. Ecological consequences of song matching: breeding success and intraspecific song mimicry in Indigo Buntings. Ecology 63:401-411.
Pröve, E. 1978. Courtship and testosterone in male Zebra Finches. Z. Tierpsychol. 48: 47–67.
Raikow, R.J. and A. H. Bledsoe. 2000. Phylogeny and Evolution of the Passerine Birds. BioScience 2000 50: 487-499.
Reid, T., R. A. Suthers, N. H. Fletcher, and W. E. Blevins. 2006. Songbirds tune their vocal tract to the fundamental frequency of their song. Proceedings of the National Academy of Science 103: 5543-5548.
Roberts, T. F., K. A. Tschida, M. E. Klein, and R. Mooney. 2010. Rapid spine stabilization and synaptic enhancement at the onset of behavioural learning. Nature 463: 948-952.
Schuppe et al. 2021. Evolutionary and Biomechanical Basis of Drumming Behavior in Woodpeckers. Frontiers in Ecology and Evolution 9: 649146.
Slabbekoorn, H. and A. den Boer-Visser. 2006. Cities change the songs of birds. Current Biology 16: 2326-2331.
Smith, G. T., E. A. Brenowitz, M. D. Beecher, and J. C. Wingfield. 1997. Seasonal Changes in Testosterone, Neural Attributes of Song Control Nuclei, and Song Structure in Wild Songbirds. Journal of Neuroscience 17:6001-6010.
Spencer, K. A., K. L. Buchanan, S. Leitner, A. R. Goldsmith, and C. K. Catchpole. 2005. Parasites affect song complexity and neural development in a songbird. Proc. Royal Sci. B 272: 2037-2043.
Templeton, C. N. and E. Greene. 2007. Nuthatches eavesdrop on variations in heterospecific chickadee mobbing alarm calls. Proceedings of the National Academy of Sciences USA 104: 5479-5482.
Templeton, C.N., E. Greene, & K. Davis. 2005. Allometry of Alarm Calls: Black-capped Chickadees Encode Information About Predator Size. Science 308:1934-1937.
Thomas, R. J., T. Székely, I. C. Cuthill, D. G. C. Harper, S. E. Newson, T. D. Frayling, and P. D. Wallis. 2002. Eye size in birds and the timing of song at dawn. Proceedings of the Royal Society B 269: 831 - 837.
Thompson, W.L. 1972. Singing behavior of the Indigo Bunting. Z. Tierpsychol. 31:39-59.
Thorpe, W. H. 1958. The learning of song patterns by birds with especial reference to the song of the Chaffinch, Fringilla coelebs. Ibis 100: 535–570.
Ward, S. and P. J. B. Slater. 2005. Heat transfer and the energetic cost of singing by Canaries Serinus canaria. Journal of Comparative Physiology A 191: 953-964.
Wilson, P. L. and S.L. Vehrencamp. 2001. A test of the deceptive mimicry hypothesis in song-sharing Song Sparrows. Anim. Behav. 62: 1197–1205.
Yamaguchi, A. 1998. A sexually dimorphic learned birdsong: the Northern Cardinal. Condor 100: 504-511.
Yamaguchi, A. 2001. Sex differences in vocal learning. Nature 411: 257-258.
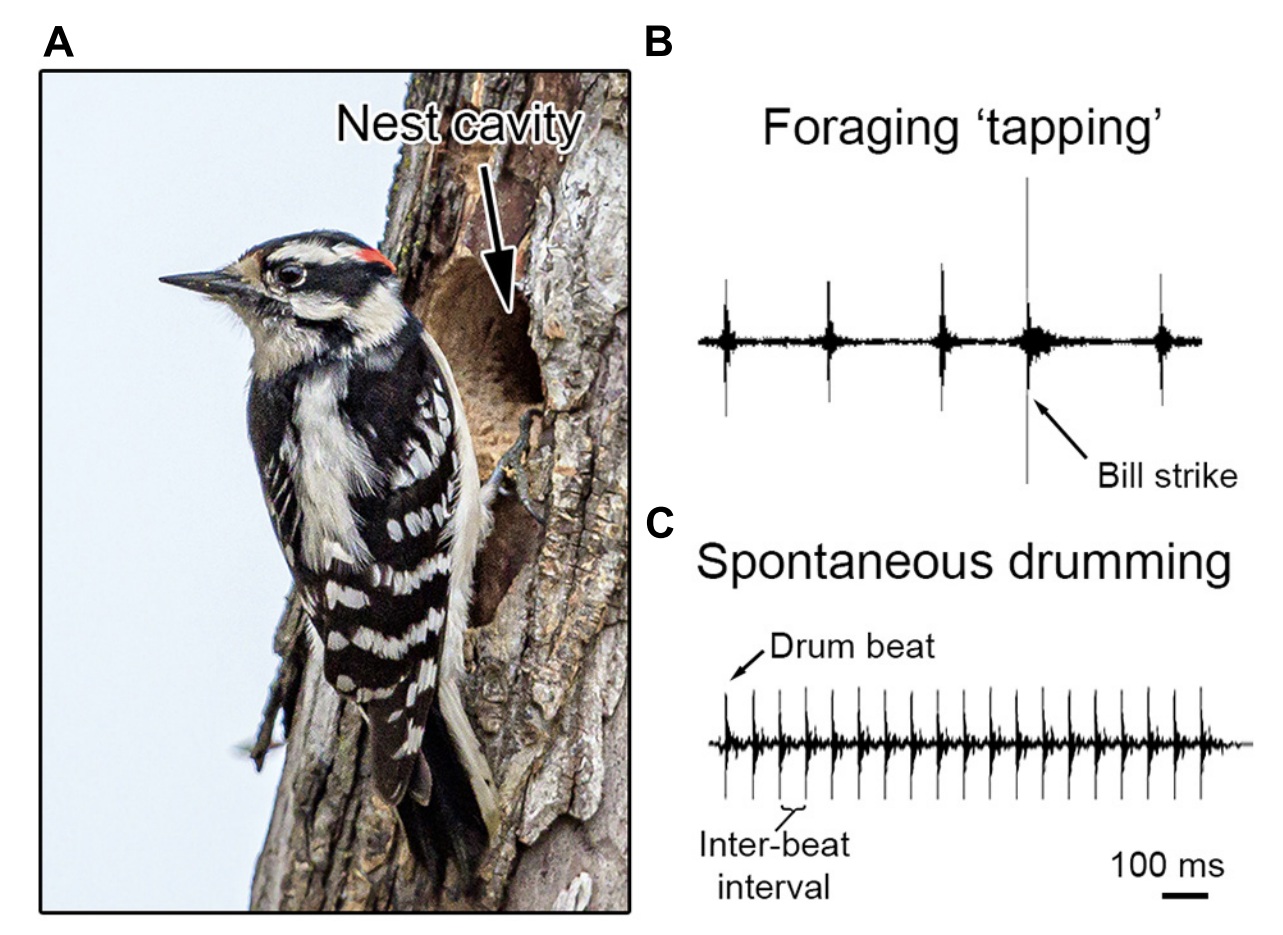
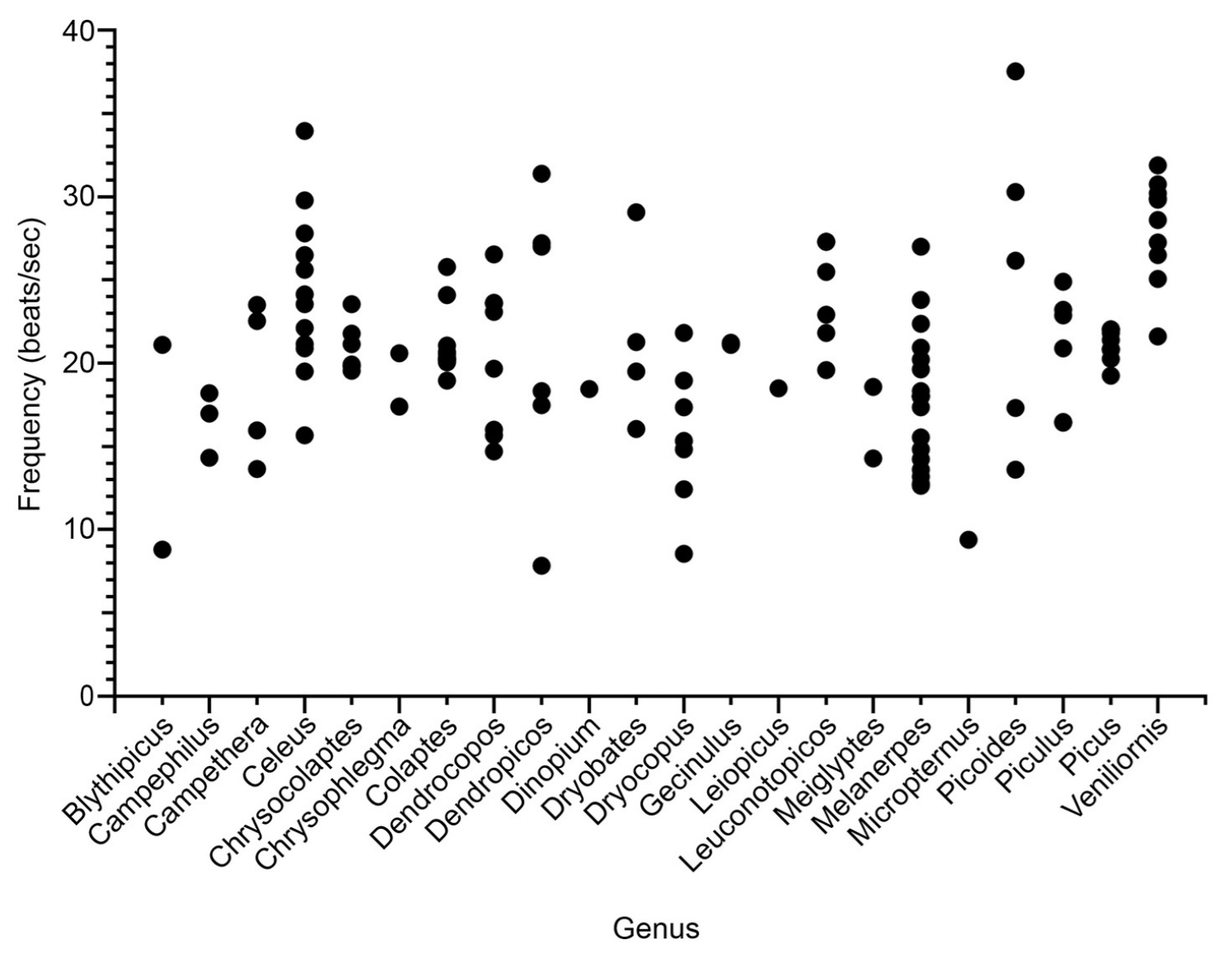
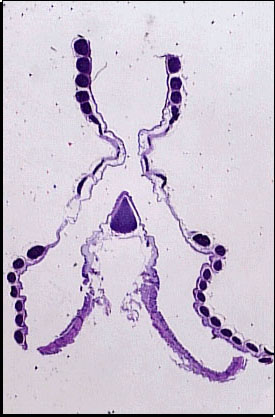
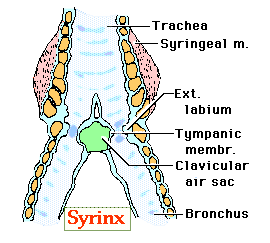
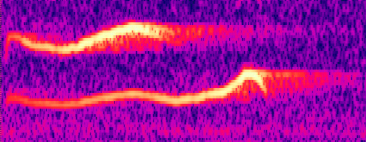
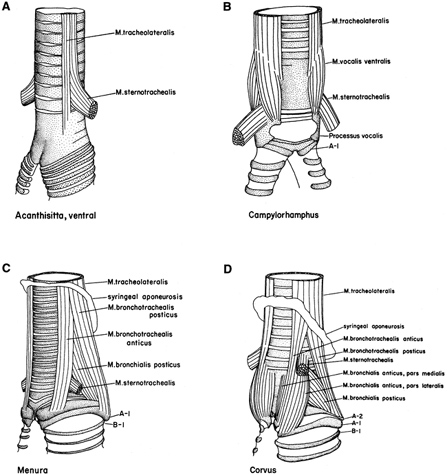

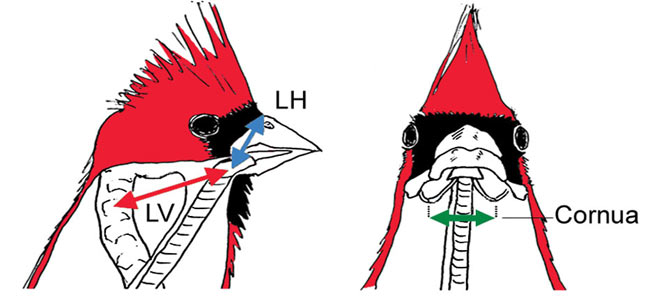
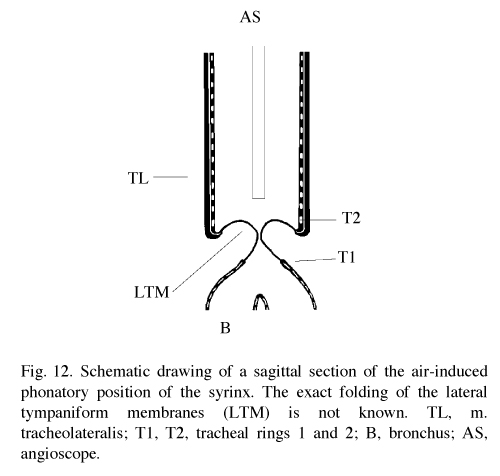
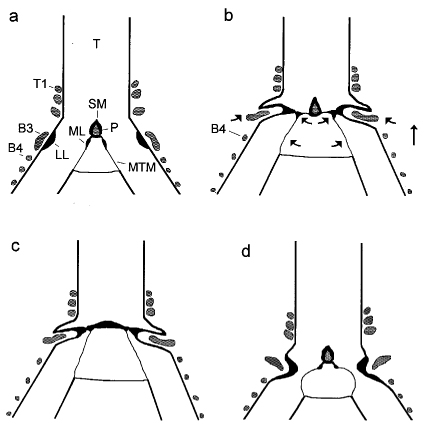

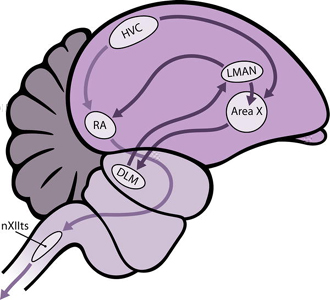
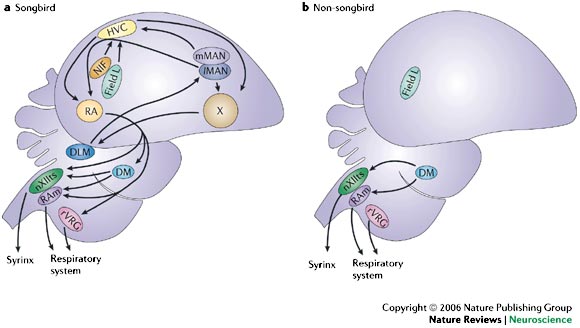

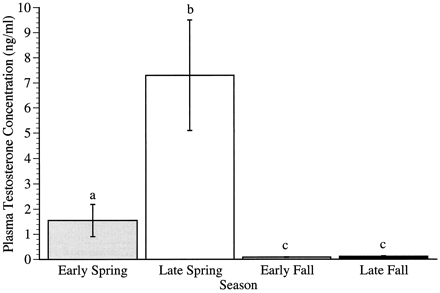
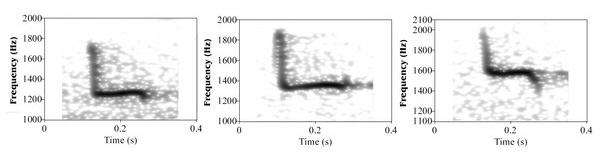

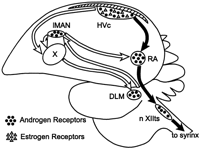
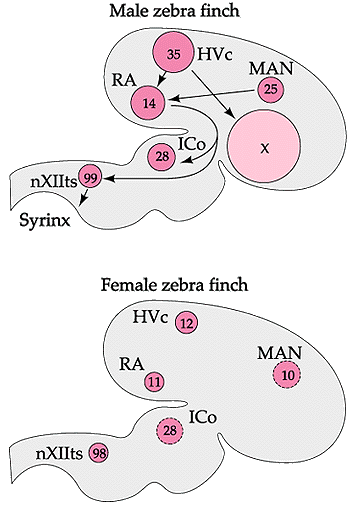
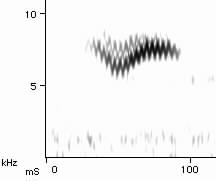
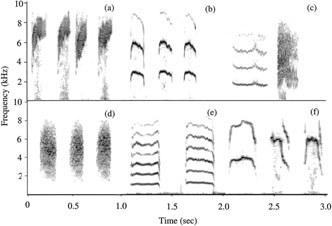 Sonograms of distress calls of six species. (a) Sooty-capped Bush-Tanager, (b) Black-capped Flycatcher, (c) Green Violet-ear, (d) Gray-breasted Wood-Wren, (e) Streak-breasted Treehunter, and (f) Yellowish Flycatcher. Each sonogram represents 1 sec of calling.
Sonograms of distress calls of six species. (a) Sooty-capped Bush-Tanager, (b) Black-capped Flycatcher, (c) Green Violet-ear, (d) Gray-breasted Wood-Wren, (e) Streak-breasted Treehunter, and (f) Yellowish Flycatcher. Each sonogram represents 1 sec of calling. 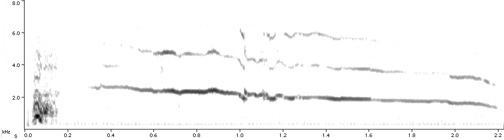
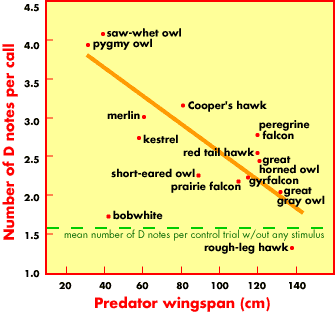

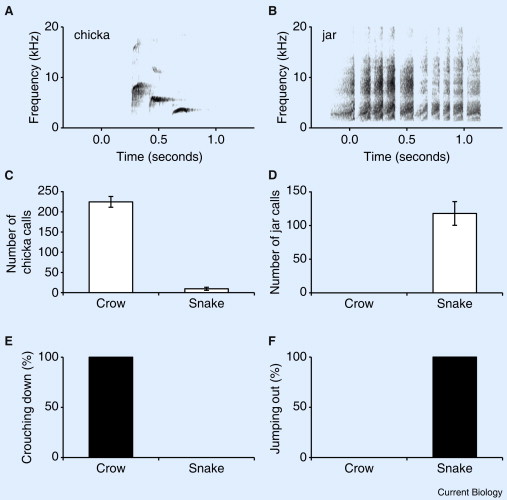

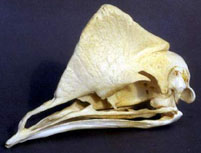
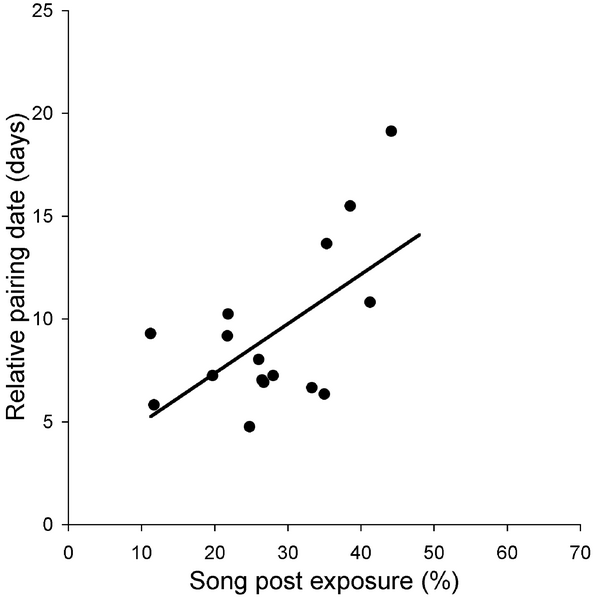
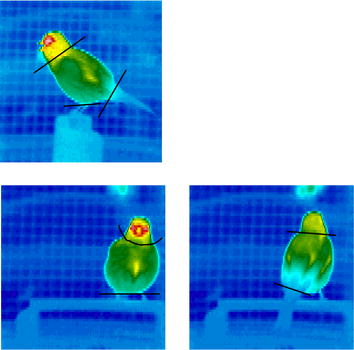
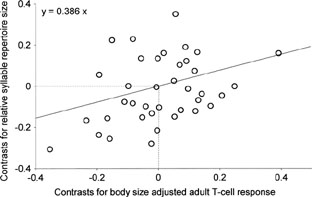
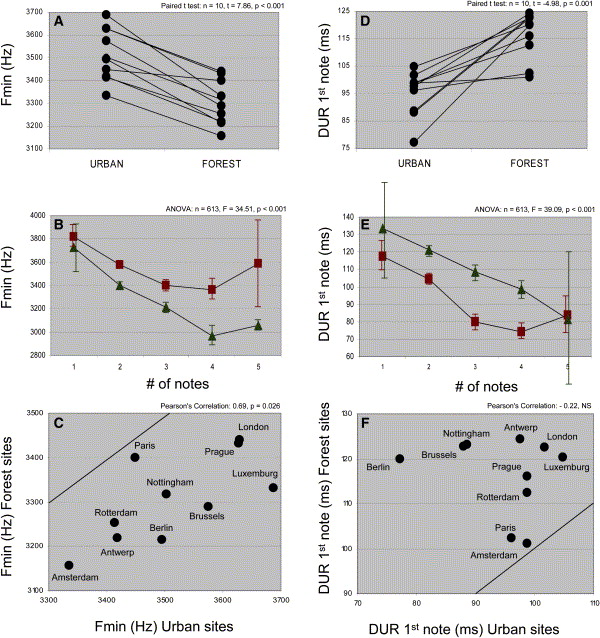


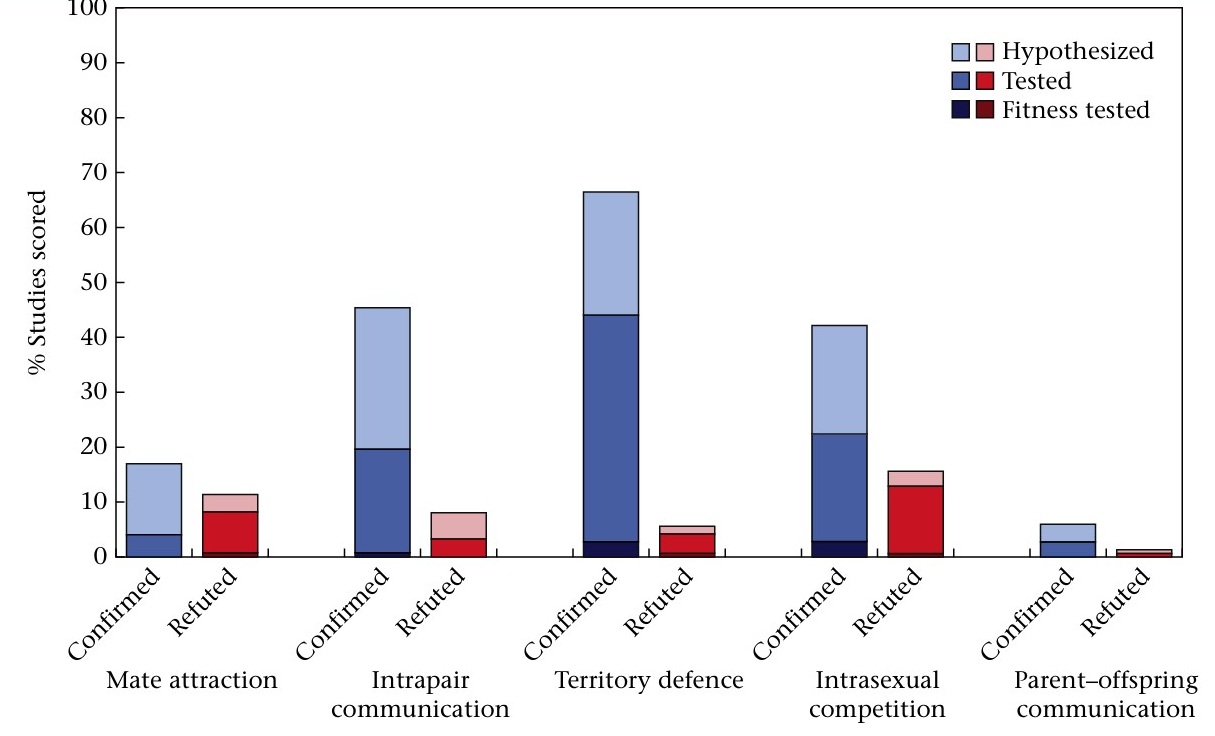


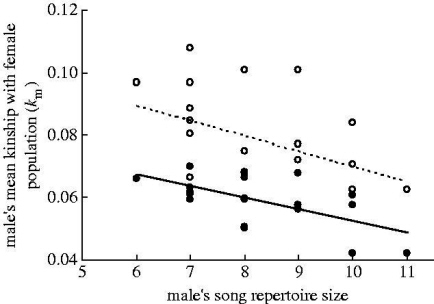
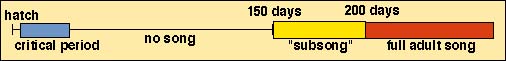
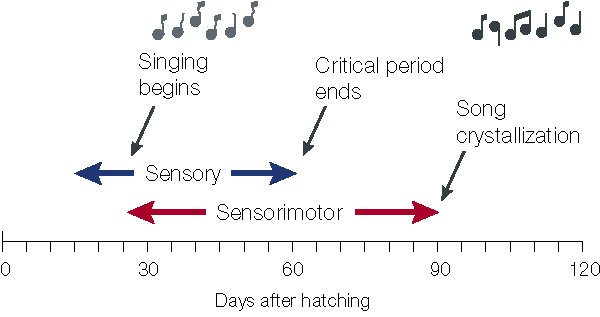


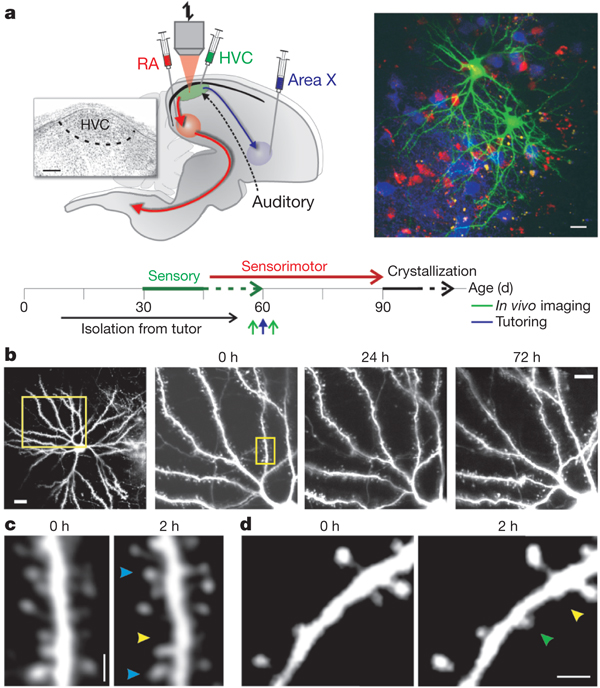

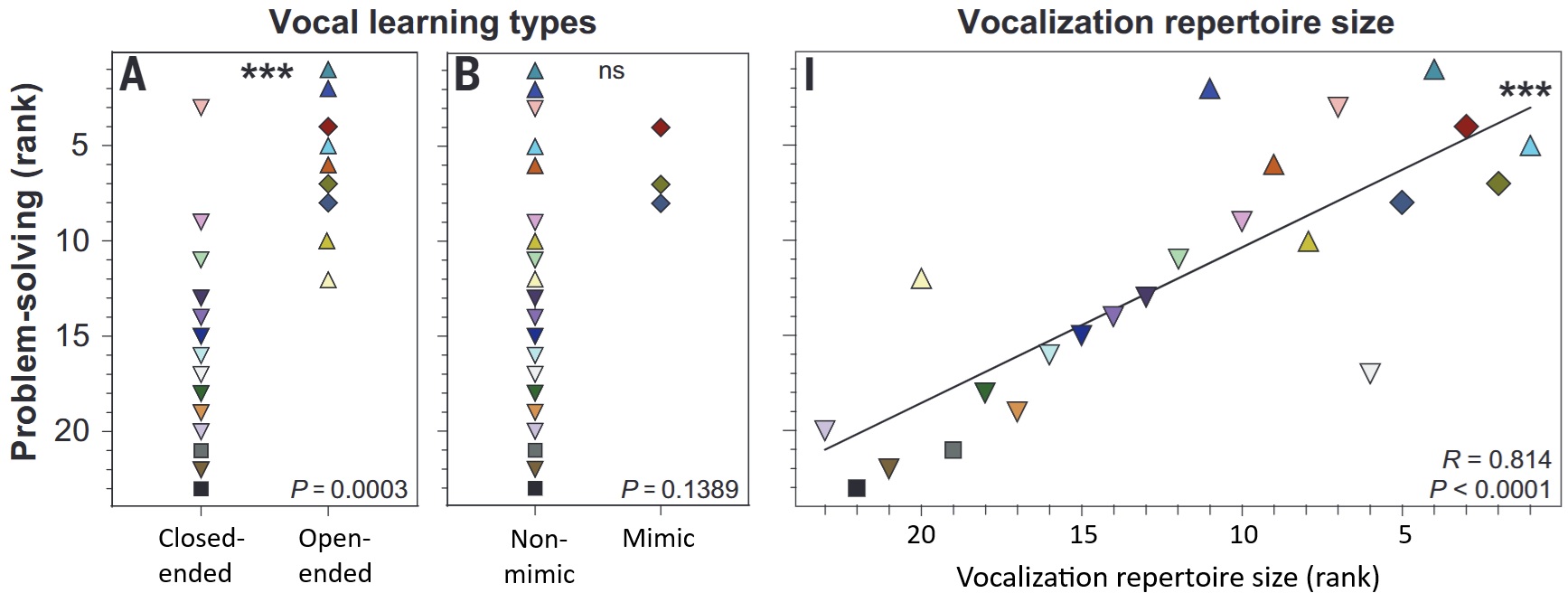

{kind=link}