Mate loss and territory proximity cause breeding dispersal in Canada Jays
Post-fledging dispersal of Purple Martins

Diagram of predator model. The first nest (solid circle at bottom) is not located within a predator's home range, and the
second nest (solid circle, upper right) would have failed on the day that the kill area contained the nest site. Powell and Frasch
(2000) simulated predation by randomly placing a predetermined density of predators
within a 100-km2 nesting area. Each predator
hunted within a home range defined by maximum movements from a central
location and a "daily kill area" (shaded boxes). Each day, each
predator moved from its central location to a point determined by x- and y-axis values randomly selected from a uniform distribution. The
predator caused nest failure if the nest was located within a fixed area surrounding the hunt location, the "daily kill area." After hunting,
the
predator
returned to the central location at the end of the day.
Nest predation and breeding dispersal -- Many types of predators depredate bird nests and thus potentially influence the spatial distribution of their prey. Powell and Frasch (2000) used a simulation model of a double-brooded songbird's nesting season to test three predictions about the selective advantage of dispersing different distances after nest predation by predators with varying home range sizes. Their results supported the predictions that (1) dispersing birds had higher success than nondispersing birds after predation of the first nest, (2) dispersing beyond the home range of the nest predator increased the success of the second nest, and (3) birds whose first nests were depredated early in the nesting cycle did better by dispersing farther than birds whose nests were depredated later in the nesting cycle. These results provide evidence that predation and predator characteristics may cause variation in adult dispersal distances during the breeding season. However, Powell and Frasch (2000) did not find an advantage for long-distance dispersal when predators with small- or medium-sized home ranges were responsible for the predation event. The critical decisions of dispersal and dispersal distance made by adult birds are complex, but it appears that predation events can create a selective advantage to disperse.
Why do individuals disperse?
Proximate vs. ultimate causation
Proximate causation:
Environmental dispersal - individuals disperse in response to environmental factors, e.g., absence of, or competition for, suitable resources or parental aggression
'Innate' dispersal - individuals are genetically predisposed to disperse
Experimental evidence for environmental dispersal:
Spanish Imperial Eagles (Ferrer 1992) - duration of pre-dispersal period for young eagles is correlated with blood levels of urea and uric acid (increased levels = reduced food intake & increased need to metabolize available proteins = poorer condition = delayed dispersal)

Spanish Imperial Eagle
Experimental evidence for innate dispersal:
Spruce Grouse (Keppie and Towers 1992) - young disperse despite removal of parents
Marsh Tits (Nilsson and Smith 1985) - young disperse despite removal of parents
Eastern Screech-Owls (Ritchison et al. 1992) - young exhibit increased locomotor activity as time of dispersal approaches
Possible causes of innate dispersal:
Eastern Screech-Owls - ontogenetic switch; time of dispersal may be correlated with condition

Western Screech-Owl
(Source: dwrcdc.nr.utah.gov/rsgis2/search/Display.asp?FlNm=otuskenn)
Corticosterone, body condition and locomotor activity: a model for dispersal in screech-owls -- Belthoff and Dufty (1998) proposed a dispersal model based on interactions among hormonal changes, body condition and social stimuli, predicting that corticosterone, an adrenal glucocorticoid known to stimulate locomotor and foraging activity, increases in blood plasma prior to dispersal through a combination of endogenous and exogenous events. This mediates the locomotor activity that underlies dispersal behaviour. Juveniles in good body condition (i.e. those with sufficient fat reserves) will disperse when corticosterone increases. Birds in poor body condition will not, but they will increase their foraging activity under the influence of corticosterone. Dispersal of siblings will reduce aggression and/or competition for food, enabling the remaining juveniles to improve their body condition and disperse. Initial studies on screech-owls, Otus asio and O. kennicottii, have produced results that are generally consistent with the model. For example, captive juvenile screech-owls showed increased locomotor activity in the weeks leading up to the time when free-living juveniles are dispersing, and activity levels declined thereafter. Peaks in corticosterone corresponded with periods of high locomotor activity (i.e. at the time of dispersal) in captive owls. Finally, field studies indicate that dominant juveniles that are presumably in better physical condition, initiated dispersal before their more subordinate siblings.
Ultimate factors responsible for dispersal:
1) Inbreeding avoidance - juveniles disperse from their natal site to prevent breeding with relatives
2) Competition for mates
3) Competition for resources (e.g., breeding territories)
Which is correct??
Most studies indicate that resource competition is an important factor, but others support mate competition or inbreeding avoidance

Predation and the cost of dispersal -- Ecologists often assume that dispersing individuals experience increased predation risk owing to increased exposure to predators while moving. To test the hypothesis that predation risk is a function of movement distance or rate of movement, Yoder et al. (2004) used radio-telemetry data collected from 193 Ruffed Grouse (Bonasa umbellus) in southeastern Ohio. They found evidence indicating that increased movement rates may increase the risk of predation for adult birds but not juveniles. They also found juvenile and adult birds inhabiting unfamiliar space were consistently at a much higher risk of predation (three to 7.5 times greater) than those in familiar space. These results indicate that although movement itself may have some effect on the risk of being preyed upon, moving through unfamiliar space has a much greater effect on risk for Ruffed Grouse. Individuals moving through unfamiliar space may suffer from decreased foraging efficiency (which also may affect energetic condition) or a decreased ability to avoid predators. A defense mechanism used by Ruffed Grouse is concealment in dense brush and undergrowth. Being in unfamiliar space may compromise this defense mechanism. Grouse moving through unfamiliar space may be more vulnerable to predation during foraging. Lack of experience in a new area may lead to difficulty locating food sources that also provide good cover. Although Yoder et al. (2004) based their analysis on distinguishing between familiar and unfamiliar space, the possibility that new territory is not only less familiar but also of lower quality (i.e., with less available cover or food sources) than is an individual's original home range cannot be ruled out. Overall, these results support the hypothesis that increased predation risk may be an important cost of dispersal for birds.
Sex-biased dispersal
Among birds, females are the predominant dispersing sex. In contrast, among
mammals, males are the predominant dispersing sex.
Reasons why dispersal may be more costly to female mammals:
Female reproductive success is limited primarily by nutritional constraints, while males are limited by number of females they can inseminate. So, females may benefit more than males from familiarity with food resources, denning sites, & neighbors (i.e., philopatry)
Because male reproductive success depends more on access to mates, they may benefit by moving to areas or groups where there are larger numbers of mates (e.g., lions & some primates)
Because intrasexual competition for mates is likely to be more intense among males in polygynous species, males are more likely to be evicted & have to disperse to find mates
What about birds?? Possible asymmetry in costs & benefits:
Avian mating systems are largely based on resource defense (males compete for & defend resources needed for successful reproduction). So, males might be more successful in establishing territories in their natal area because familiarity may permit higher feeding rates & reduced predation rates.
Females might benefit from the potential to choose between the resources of different mates &, if inbreeding is costly, females rather than males should disperse.
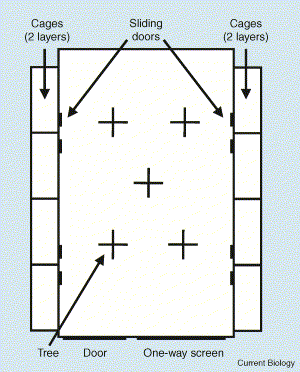
Plan of the ‘novel environment’ room in which the exploratory behavior of wild-caught Great Tits was tested by Dingemanse et al. (2003).
The room has dimensions 4.0 × 2.4 × 2.3 m. Along each 4-m wall were eight sliding doors, in two rows of four above each other, connecting the holding
cages to
the room. The front 2.4m wall had a 0.9 × 2.0 m door at the left side and a 1.1 × 0.16 m one-way screen through which the birds could be observed.
The room contained five artificial trees made of wood with a trunk of 4 × 4 cm and a height of 1.5 m. Each tree had four cylindrical branches 20 cm long. The
upper two branches (5 cm below the top) were on opposite sides of the trunk, perpendicular to the lower branches (25 cm below the top). Birds entered
the room through one of the sliding doors (Dall 2004).
Natal dispersal and personalities in Great Tits (Parus major) -- Dispersal is a major determinant of the dynamics and genetic structure of populations, and its consequences depend not only on average dispersal rates and distances, but also on the characteristics of dispersing and philopatric individuals. Dingemanse et al. (2003) investigated whether natal dispersal correlated with a predisposed behavioural trait: exploratory behaviour in novel environments. Wild Great Tits were caught in their natural habitat, tested the following morning in the laboratory using an open field test and released at the capture site. Natal dispersal correlated positively with parental and individual exploratory behaviour, using three independent datasets. First, fast-exploring parents had offspring that dispersed furthest. Second, immigrants were faster explorers than locally born birds. Third, post-fledging movements, comprising a major proportion of the variation in natal dispersal distances, were greater for fast females than for slow females. These findings suggest that parental behaviour influenced offspring natal dispersal either via parental behaviour per se (e.g. via post-fledging care) or by affecting the phenotype of their offspring (e.g. via their genes). Because this personality trait has a genetic basis, these results imply that genotypes differ in their dispersal distances. Therefore, the described patterns have profound consequences for the genetic composition of populations.
Natal dispersal of Canada Jays

Natal dispersal distances of four species of birds in Great Britain (European Blackbird, Turdus merula;
Reed Warbler, Acrocephalus scirpaceus; Lesser Black-backed Gull, Larus fuscus; and Woodpigeon, Columba palumbus)
(Paradis et al. 1998).
How far do young disperse?
Median dispersal distances for most birds is less than 10 home range diameters. Why?
Assume:
individuals do not share ranges (or territories)
adults (residents) are dominant to dispersing young
So, dispersing young settle on the first uncontested site they find & uncontested sites arise through the death of adults.
But, if true, then why do some young disperse substantial distances & what about young that become floaters & do not obtain and defend a site? More research is needed.
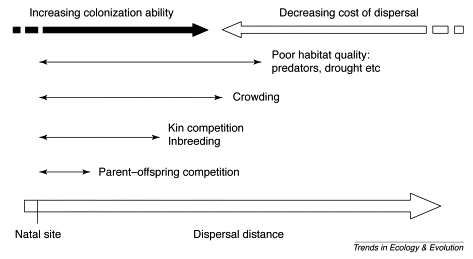
The evolution of different dispersal distances might involve multiple selective forces operating
at different spatial scales (Ferriere et al. 2000).
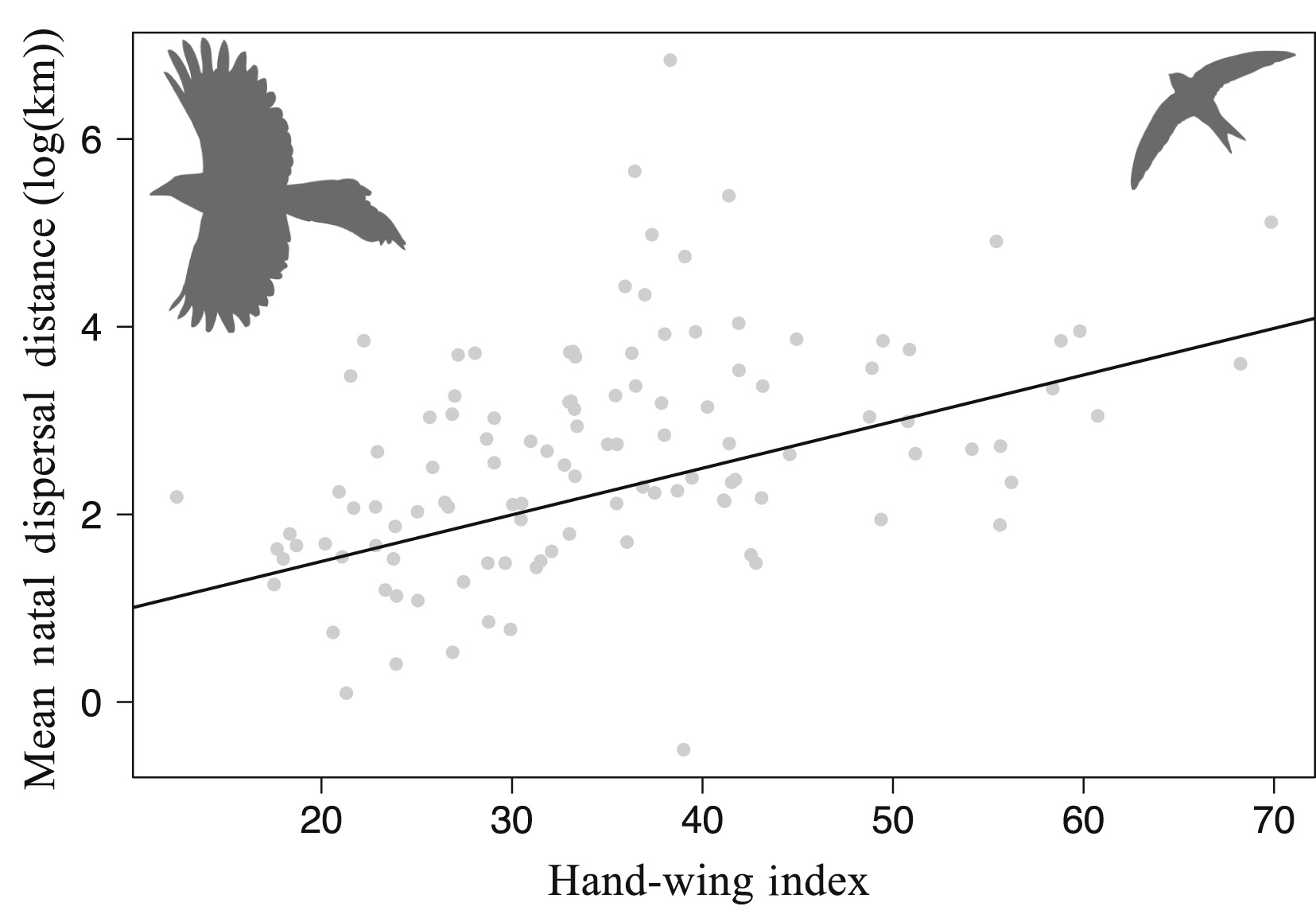
Weeks et al. (2022) found that hand-wing Index (HWI; a morphological proxy for wing aspect ratio) is positively
related to mean natal dispersal distance. Each point represents the mean HWI and dispersal distance for 114 species of
birds. Natal dispersal involves the departure of individuals from their natal sites. This dispersal may be influenced by
various factors, including species biology, biotic interactions and geographical barriers. However, the results of this study suggest that
flying ability is clearly a critical factor in bird dispersal, i.e., species adapted for efficient flight disperse further, perhaps because they are
more mobile and thus are capable of crossing gaps in suitable habitat. As a result, they are able to travel longer distances on average during
the movement phase.
Natal dispersal of Florida Scrub-Jays

Floater home ranges and prospective behavior of European Starlings -- In many bird species, floaters are present on the breeding grounds in one or more years before they breed. There is increasing evidence that they have specific home ranges in which they search for information about current and future breeding opportunities. Tobler and Smith (2004) investigated the role of prospecting in a migratory European Starling (Sturnus vulgaris) population. Radio-tracking showed that male starling floaters use specific home range areas during the breeding period. Nest-box observations demonstrated that non-parental nest intrusion is common in the starling and that it is significantly more frequent during the nestling than during the incubation period. In addition, small groups of nest boxes were more likely to be occupied by starlings if they had been put up during the preceding breeding season. These results suggest that floaters try to acquire information about local breeding communities. One specific type of information may be the location of potential breeding sites.
Number of fledglings produced by birds that grew up in a parasite-free nest (open circles and dashed line) or a nest infested with ectoparasitic hen fleas (solid circles and solid line) in relation to their natal dispersal distance. Residuals of the model without the interaction between parasite treatment and natal dispersal distance are shown. |
Natal dispersal distance of recruits originating from testosterone- |
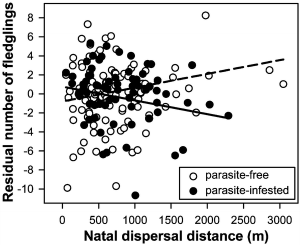
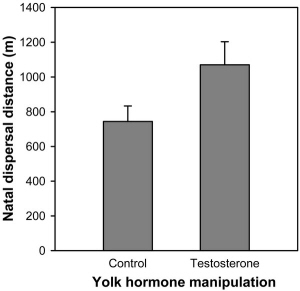
Maternal modulation of natal dispersal in Great Tits -- The decision of how far to disperse from the natal territory has profound and long-lasting consequences for young animals, yet the optimal dispersal behavior often depends on environmental factors that are difficult or impossible to assess by inexperienced juveniles. Natural selection thus favors mechanisms that allow the adaptive and flexible adjustment of the offspring's dispersal behavior by their parents via either paternal or maternal effects. Tschirren et al. (2007) showed that different dispersal strategies maximize the reproductive success of young Great Tits (Parus major) originating from a parasite-infested or a parasite-free nest and demonstrate that differential transfer of maternal yolk androgens in response to parasitism can result in a modification of the offspring's dispersal behavior that appears adaptive. These results demonstrate that prenatal maternal effects are an important yet so far neglected determinant of natal dispersal and highlights the potential importance of maternal effects in mediating coevolutionary processes in host-parasite systems.
Literature cited:
Belthoff, J. R. and A. M. Dufty, Jr. 1998. Corticosterone, body condition and locomotor activity: a model for dispersal in screech-owls. Animal Behaviour 55: 405-415.
Dall, S. R. X. 2004. Behavioural Biology: Fortune Favours Bold and Shy Personalities.Current Biology 14: R470-R472.
Dingemanse, N. J., C. Both, A. J. van Noordwijk, A. L. Rutten , P. J. Drent. 2003. Natal dispersal and personalities in Great Tits (Parus major). Proceedings of the Royal Society B 270: 741-747.
Ferrer, M. 1992. Natal dispersal in relation to nutritional
condition in Spanish Imperial Eagles. Ornis Scand. 23:104-107.
Ferriere, R., J. R. Belthoff, I. Olivieri, and S. Krackow. 2000. Evolving dispersal: where to go next? Trends in Ecology and Evolution 15: 5-7.
Keppie, D.M. and J. Towers. 1992. A test on social behavior
as a cause of dispersal of Spruce Grouse. Behav. Ecol. Sociobiol. 30:343-346.
Nilsson, J.A. and H.G. Smith. 1985. Early fledgling mortality
and the timing of juvenile dispersal in the Marsh Tit Parus palustris.
Ornis Scand. 16:293-298.
Paradis, E., S. R. Baillie, W. J. Sutherland, and R. D. Gregory. 1998. Patterns of natal and breeding dispersal in birds. Journal of Animal Ecology 67: 518-536.
Powell, L. A., and L. L. Frasch. 2000. Can nest predation and predator type explain variation in dispersal of adult birds during the breeding season? Behavioral Ecology 11: 437-443.
Ritchison, G., J. R. Belthoff, and E.J. Sparks. 1992. Dispersal
restlessness: evidence for innate dispersal by juvenile Eastern Screech-Owls?
Anim. Behav. 43:57-65.
Tobler, M. and H. G. Smith. 2004. Specific floater home ranges and prospective behaviour in the European Starling, Sturnus vulgaris. Naturwissenschaften 91: 85-89.
Tschirren, B., P. S. Fitze, and H. Richner. 2007. Maternal modulation of natal dispersal in a passerine bird: an adaptive strategy to cope with parasitism? American Naturalist 169: 87-93.
Weeks et al. 2022. Morphological adaptations linked to flight efficiency and aerial lifestyle determine natal dispersal distance in birds. Functional Ecology 36: 1681-1689.
Yoder, J. M., E. A. Marschall , and D. A. Swanson. 2004. The cost of dispersal: predation as a function of movement and site familiarity in Ruffed Grouse.
Behav. Ecol. 15: 469-476.