Bird Biogeography |
 |
Bird Biogeography |
|
Currently, taxonomists recognize 44 orders, 256 families, 2396 genera, and well over 10,000 species of birds (AvianLexiconAtlas: A database of descriptive categories of English-language bird names around the world). Birds occupy all continents and habitats but, of course, some continents & some habitats have more species than others.
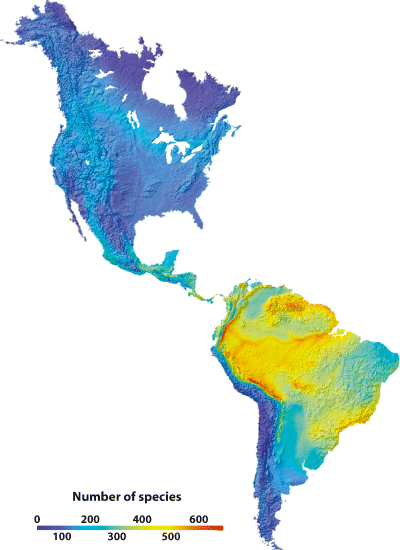
Bird species richness in the Western Hemisphere (Hawkins et al. 2006).
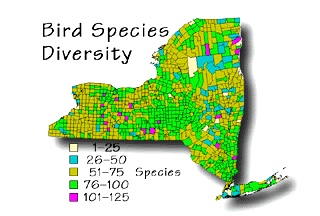
Bird species diversity in New York state
How are birds currently distributed throughout the planet and what factors have contributed to this distribution? In part, the current distribution of birds has been influenced by historical events. Birds belonging to various lineages have been found in Cretaceous deposits of Asia, Europe, and North and South America. Prominent and well-known Cretaceous bird taxa included the Enantiornithes, a fairly diverse group of birds, mostly flying forms; Hesperornithiformes, toothed birds (see drawing below) which were mostly flightless swimmers; and Ichthyornithiformes, toothed flying birds that probably fed on fish. These taxa are extinct today, but by the close of the Cretaceous, representatives of several modern bird taxa were sharing the skies with these extinct birds. A birdwatcher 65 million years ago could have seen relatives of today's loons, geese and ducks, albatrosses and petrels, and gulls and shorebirds, and possibly other familiar birds as well.

Loon-like toothed bird Hesperornis regalis
swims
through the Cretaceous sea
(Painting by Dan Varner; source: www.dmr.nd.gov/ndfossil/Education/trunks/Trunk%20030609.html)
What does the fossil record reveal concerning the present-day distribution of birds? The study of historical bird geography is difficult because of the many variables involved: evolution, climatic change, shifting vegetation, weather disasters, epizootics, rapid dispersal, ecological adaptation, competition between species, and barriers (Welty & Baptista 1988). However, it is known that (Proctor and Lynch 1993):

Cretaceous enantiornithines. Top: male Feitianius, center left: female Feitianius, center: Orienantius, center right: Sulcavis,
Bottom left: Avimaia, bottom center: Falcatakely, bottom right: Longipteryx. Scale bar: 5 cm (From: O'Connor 2022).
The Enantiornithes are an extinct lineage of birds. e, membranes that hold the developing embryo of present-day birds in place.
Available information indicates that enantiornithines were highly precocial, hatching with a well ossified skeleton and fully developed remiges, presumably fully independent and capable of flight within 24 hours after hatching.
Fossilized melanosomes (organelles containing the pigment melanin found in noAerodynamic analysis suggests that Early Cretaceous enantiornithines used intermittent flapping flight styles like bounding (bursts of flapping punctuated by short intervals where the wings are folded against the body) and flap-gliding. However, these flight styles are limited to small birds with body masses of less than 300 grams. The larger enantiornithines that evolved in the Late Cretaceous must have used different flight styles, although the fragmentary nature of specimens from younger deposits prevents precise analysis.
Enantiornithines were polyphylodont, meaning their teeth were being continuously replaced throughout their lifetime. Tooth shapes vary enormously among enantiornithines, suggesting a wide diversity of diets.
Fossils have revealed that female enantiornithines had a single functional ovary and oviduct and utilized medullary bone as a calcium reservoir during eggshell production. The structure of their nests, with the eggs partially embedded in sediment, suggests that enantiornithines did not rotate their eggs during development, which suggests the absence of chalazan-white feathers) in some specimens have so far only revealed eumelanosomes (elongate melanosomes that produce black and grey coloration). However, phaeomelanosomes (rounder melanosomes that produce a reddish-brown color) and structural color (created through light interference produced by the spongy nanostructure of some feathers) were likely also present in enantiornithines. Plumage color patterns are detectible in specimens preserved in amber, revealing a diversity of wing patterns consisting of dark regions traversed by pale bands and spots.
This extinct group of birds dominated terrestrial environments from 130 to 66 million years ago. With approximately 90 known genera, they account for more than half of the known diversity of Mesozoic birds. Despite apparently out-competing birds more closely related to present-day species in most Cretaceous continental environments, enantiornithines mysteriously went extinct at the end of the Cretaceous (From: O'Connor 2022).
| Fossil
Evidence for the Extant Avian Radiation in the Cretaceous -- Clarke
et al. (2005) provide apparent evidence that cousins of living birds
coexisted
with dinosaurs more than 65 million years ago. Information from a new
species
called Vegavis
iaai indicates that these birds lived in the Cretaceous and
must
have survived the Cretaceous/Tertiary (K/T) mass extinction event that
included the disappearance of all other dinosaurs. Analysis of the
fossil,
discovered in Antarctica in 1992, revealed a new species in the
group
Anseriformes, which includes ducks and geese. The question of whether
relatives
of living birds co-existed with non-bird dinosaurs has evoked
controversy.
Some investigators, using “molecular clock” models and DNA
sequence
data as well as the distribution of living birds, have concluded that
relatives
of living birds must have existed alongside non-avian dinosaurs and
survived the mass extinction of dinosaurs at the K/T boundary. Others believe
such
data are unreliable, that the fossil record shows no evidence of living
bird lineages in the Cretaceous, and that relatives of today’s birds
evolved
after the K/T boundary.
“We have more data than ever to propose at least the beginnings of the radiation of all living birds in the Cretaceous,” Clarke says. “We now know that duck and chicken relatives coexisted with non-avian dinosaurs. This does not mean that today’s chicken and duck species lived with non-avian dinosaurs, but that the evolutionary lineages leading to today’s duck and chicken species did.” |

Skrepnick shows Vegavis in the foreground with a duckbill dinosaur in the background (Source: www.ncsu.edu). |
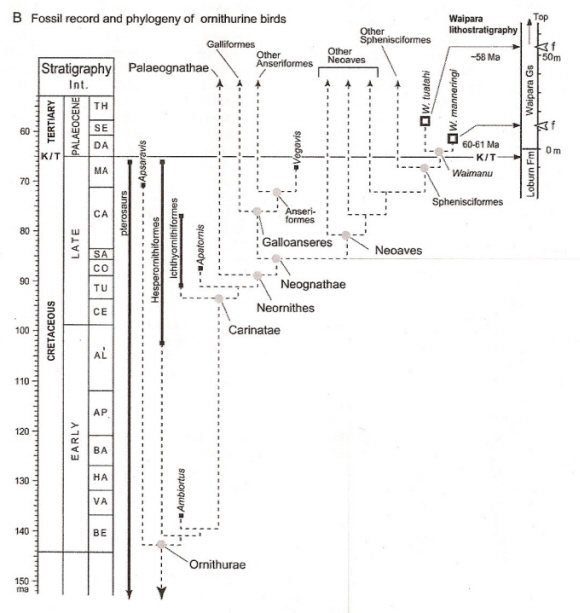
Fossil record and phylogeny of ornithurine birds (ornithurine = bird-like, as opposed to sauriurine = reptile-like, e.g, Archaeopteryx).
Solid lines show geological ranges of taxa with first and last occurrences shown by squares.
Dashed lines show postulated phylogeny compiled from the literature (Slack et al. 2006).
Calibrating avian evolution -- Slack et al. (2006) described the earliest penguin fossils and analyzed complete mitochondrial genomes from an albatross, a petrel,
and a loon. The penguin fossils were from a Paleocene (early Tertiary) formation just above a well-known Cretaceous/Tertiary boundary site.
The fossils, in a new genus (Waimanu), provide a lower estimate of 61–62 Ma for the divergence between penguins and other birds and thus
establish a reliable calibration point for avian evolution. Combining fossil calibration points, DNA sequences, maximum likelihood, and Bayesian
analysis, the penguin calibrations imply a radiation of modern (crown group) birds in the Late Cretaceous. This includes a conservative estimate that
modern sea and shorebird lineages diverged at least by the Late Cretaceous about 74 million years ago. It is clear that modern birds
from at least the latest Cretaceous lived at the same time as archaic birds including Hesperornis, Ichthyornis, and the diverse Enantiornithiformes.
Pterosaurs also coexisted with early birds. Additional fossils and molecular data are still required to help understand the role of biotic interactions
in the evolution of Late Cretaceous birds and thus to test that the mechanisms of microevolution are sufficient to explain macroevolution.
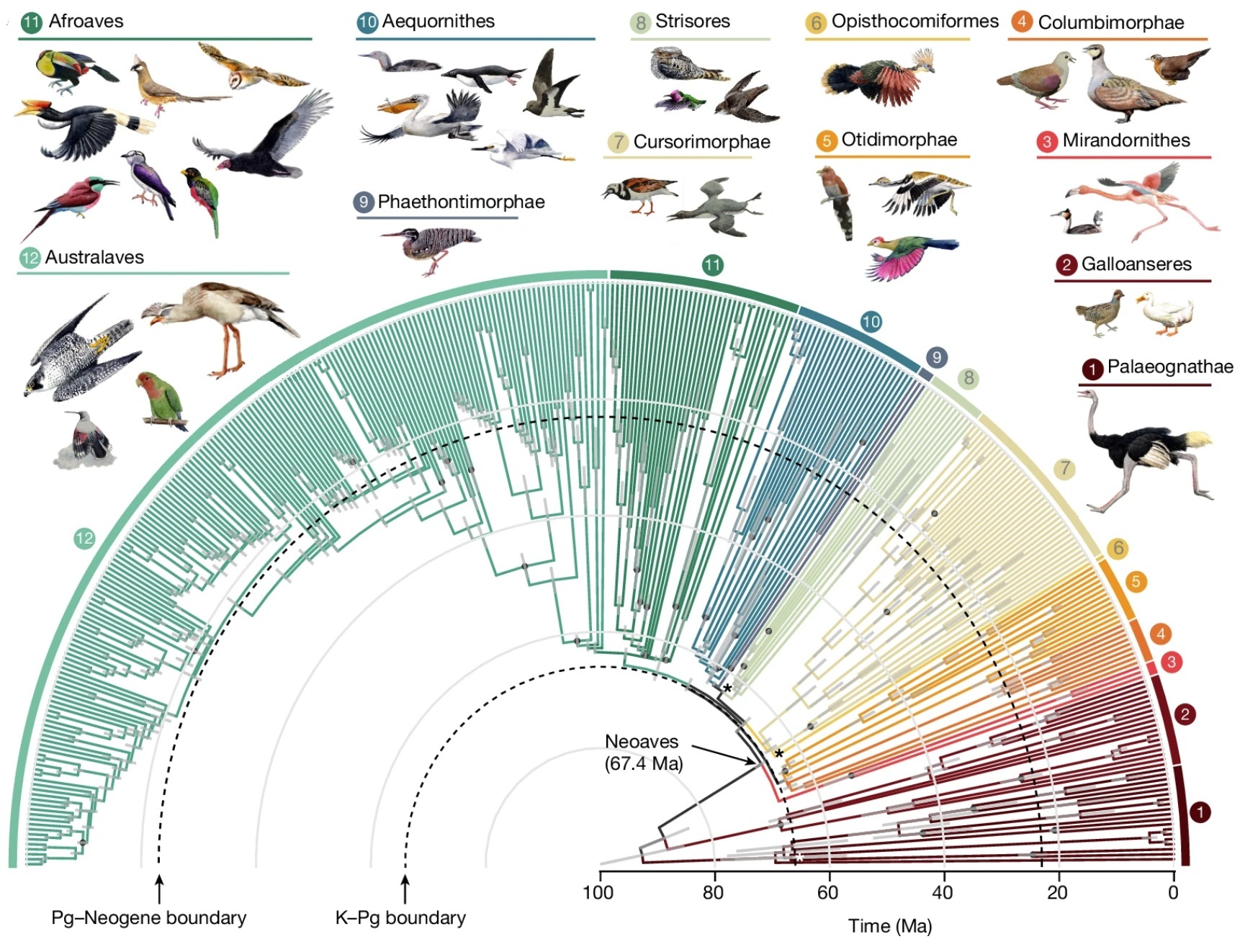
Time tree of bird evolution. Gray bars represent 95% credible intervals for age estimation; dots indicate nodes with fossil calibrations;
asterisks mark the three branches lacking full support.
Relationships and divergence times for 363 bird species. Modern birds comprise three major groups: ratites and tinamous (Palaeognathae), landfowl and waterfowl (Galloanseres) and all other living birds (Neoaves). Palaeognathae and Galloanseres appeared well before the end-Cretaceous extinction event (K-Pg boundary). The early Neoaves experienced rapid diversification into at least ten major clades. Based on their analysis, Stiller et al. (2024) suggest that the early divergences in Neoaves were tightly associated with the KPg boundary, with only two divergences before the boundary: Mirandornithes (grebes and flamingos} diverged from the remaining Neoaves 67.4 Ma and Columbaves (doves, sandgrouse and mesites) diverged 66.5 Ma. All subsequent divergences postdate the boundary. The most recent common ancestor of Passeriformes (Afroaves and Australaves in the figure above) was dated to 50.7 Ma, similar to results of other studies (4753 Ma). The split between Tyranni (Suboscines) and Passeri (Oscines) was estimated at 47.3 Ma.
The distribution of some groups of birds may have been influenced by continental drift (Cracraft 1974). Some examples of taxa where evidence indicates that intercontinental connections influenced present-day distribution include penguins and parrots:
Appearance: Primarily black-and-white plumage, often with colorful bills (like puffins) during the breeding season.
Behavior: They are deep divers, pursuing fish underwater for food. They have fast, whirring flight.
Habitats: Breed in coastal colonies on cliffs or rocky islands and spend winters at sea.
Breeding: Breeding sites vary from temperate forests to rocky, glacial outcrops.
Some Alcidae species - The family is divided into several genera, including:
Puffins (Fratercula, Cerorhinca): Atlantic Puffin (F. arctica), Tufted Puffin (F. cirrhata), Horned Puffin (F. corniculata), and Rhinoceros Auklet (C. monocerata).
Murres/Guillemots (Uria, Cepphus): Common Murre (U. aalge), Thick-billed Murre (U. lomvia), and Black Guillemot (C. grylle).
Auklets (Aethia, Ptychoramphus): Crested Auklet (A. cristatella), Least Auklet (A. pusilla), Parakeet Auklet (A. psittacula), and Cassin's Auklet (P. aleuticus).
Murrelets & Razorbill (Synthliboramphus, Brachyramphus, Alca): Razorbill (Alca torda), Marbled Murrelet (B. marmoratus), and Ancient Murrelet (S. antiquus).
Dovekie (Alle alle): The smallest Atlantic alcid.
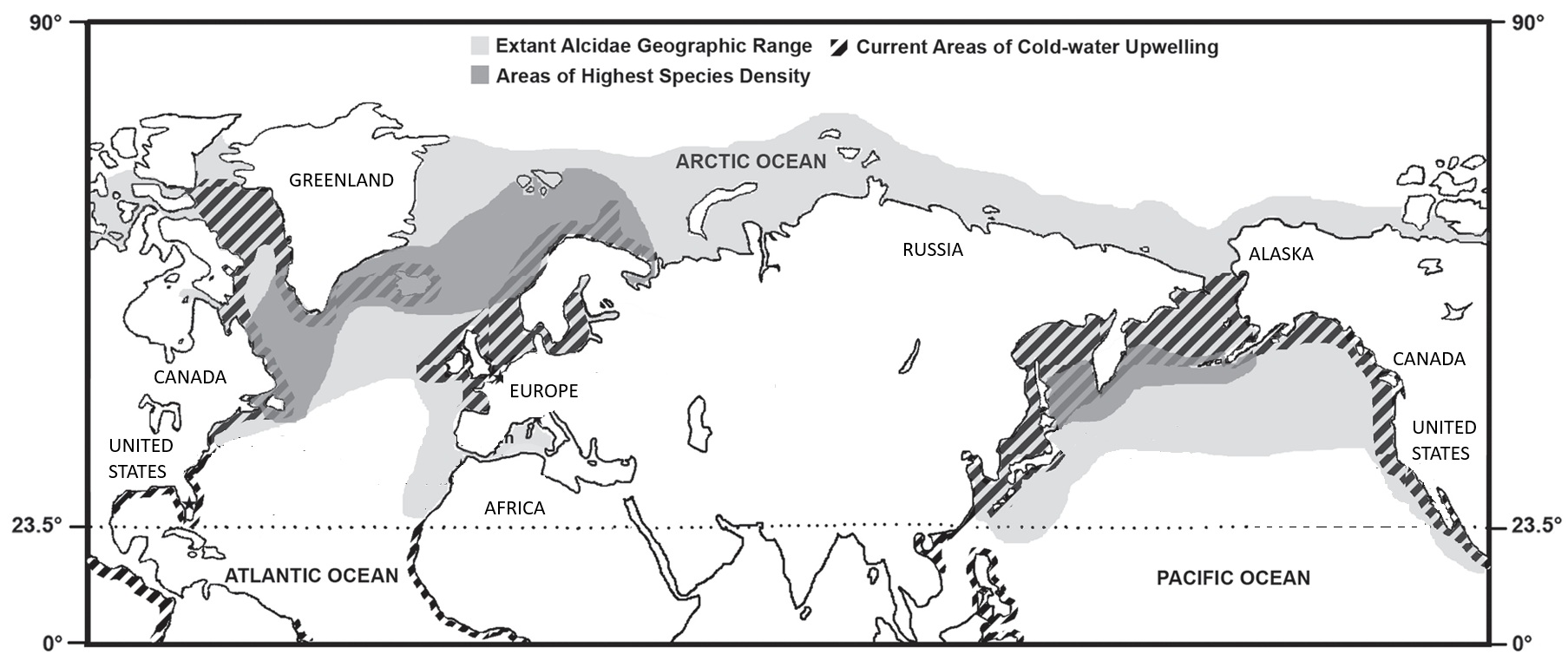
Geographic distribution of present-day alcids (light gray), the areas of highest species diversity (dark gray), and areas of modern cold-water upwelling (diagonal lines).
Note that the range of alcids extends south of the Tropic of Cancer (From: Smith and Clarke 2014).
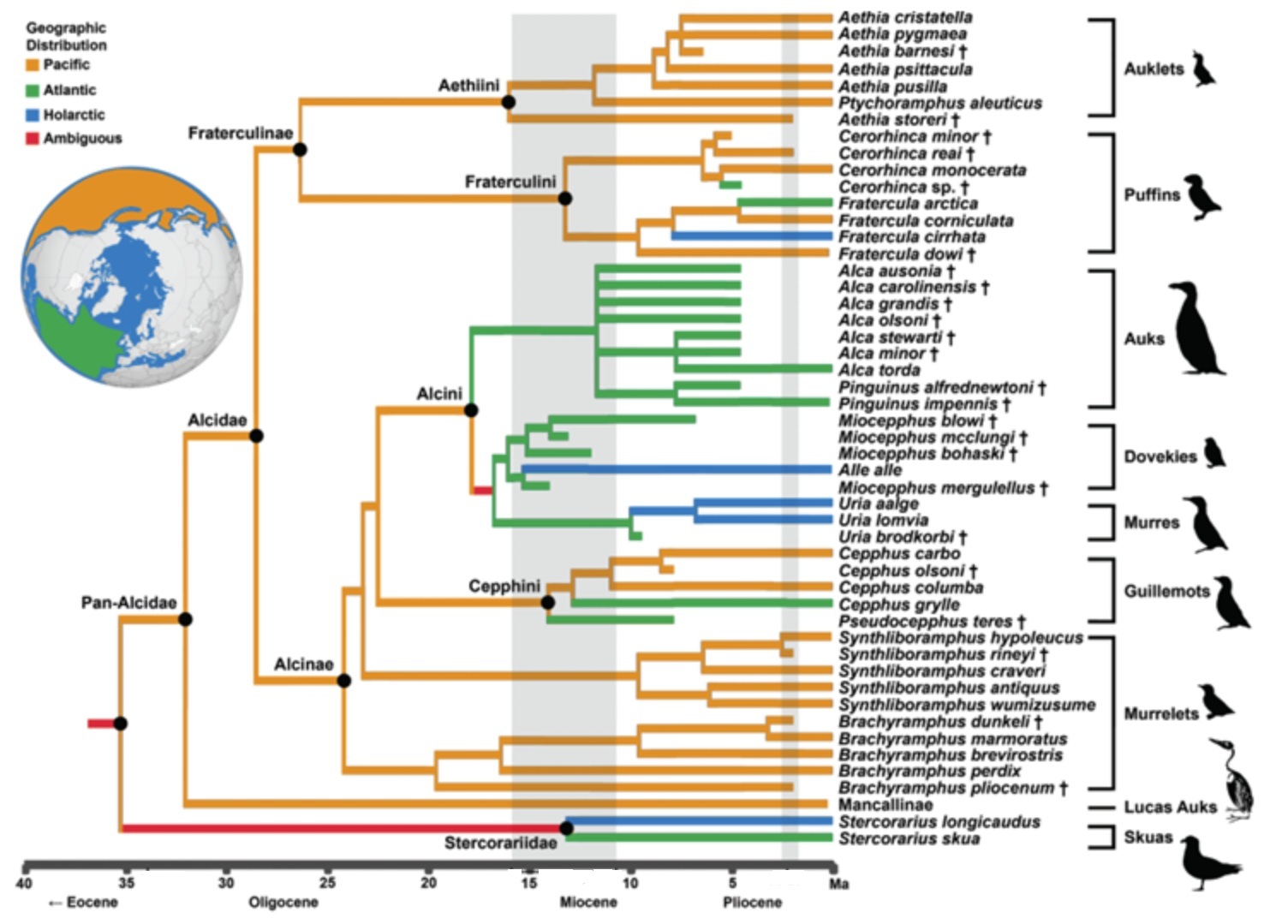
Phylogeny, divergence time estimates, and ancestral areas species in the family Alcidae (including 28 extinct species).
Cross symbol = extinct species (From: Smith and Clarke 2014).
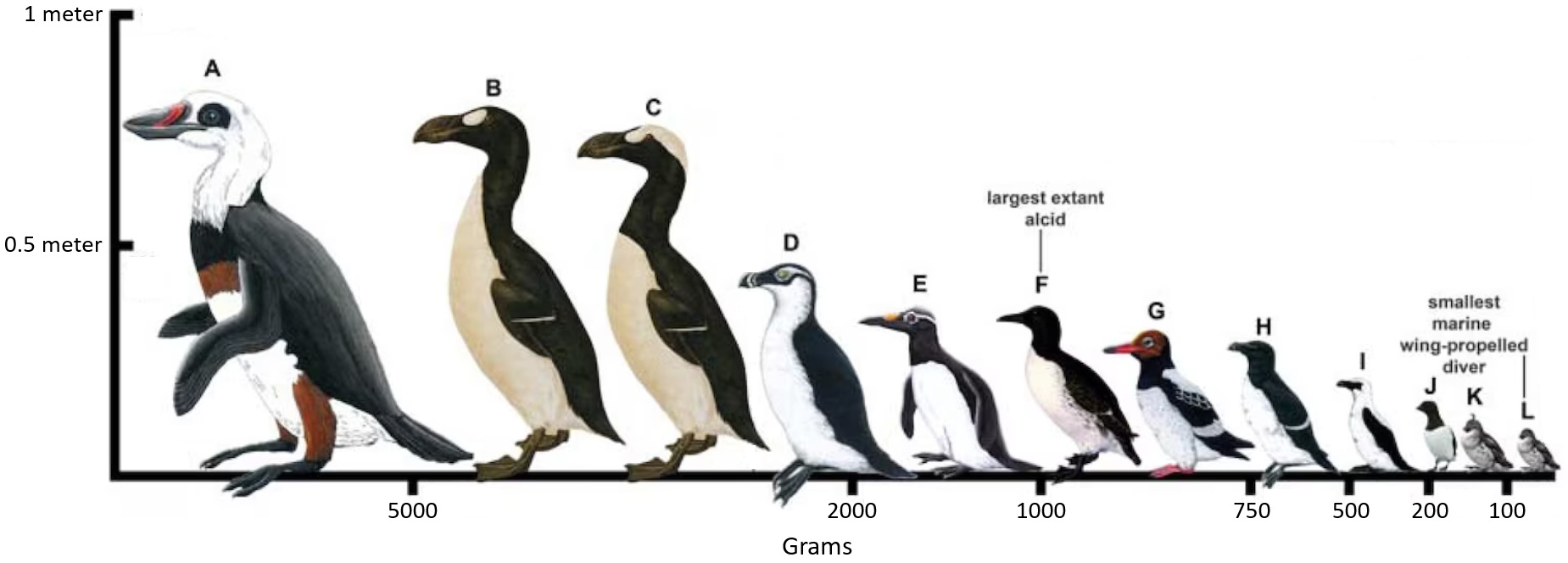
Selected species of alcids demonstrating the range of body sizes. (A) Miomancalla howardae - E, (B) Great Auk (Pinguinus impennis) - E,
(C) Pinguinus alfrednewtoni - E, (D) Alca stewarti - E, (E) Mancalla vegrandis - E, (F) Common Murre (Uria aalge, (G) Miocepphus blowi - E,
(H) Razorbill (Alca torda), (I) Alca minor - E, (J) Little Auk (Alle alle), (K) Aethia storeri - E, and (L) Least Auklet (Aethia pusilla).
E = extinct (From: Smith 2016).

Global distribution of penguins
(Source: https://penguincabin.herokuapp.com/map)
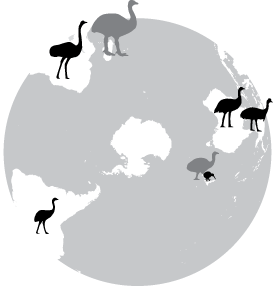
For many years, the consensus has been that the ancestors of the ratite (flightless) birds may have originated in the ancient super-continent of Gondwanaland, then became widely separated as South America, Africa, and Australia drifted off to their present locations during the Mesozoic. There they evolved into their present-day forms: ostrich, rheas, emu, cassowaries, kiwis, and moas. However, recent evidence indicates that this scenario is likely incorrect:
The prevailing view has been that ratites are monophyletic, with the flighted tinamous as their sister group, suggesting a single loss of flight in the common ancestry of ratites. However, Harshman et al. (2008) conducted phylogenetic analyses of 20 unlinked nuclear genes and found a genome-wide signal that unequivocally places tinamous within ratites, making ratites polyphyletic and suggesting multiple losses of flight. Phenomena that can mislead phylogenetic analyses (e.g., long branch attraction, base compositional bias, discordance between gene trees and species trees, and sequence alignment errors) were eliminated as explanations for this result. The most plausible hypothesis requires at least three losses of flight and explains the many morphological and behavioral similarities among ratites by parallel or convergent evolution. Finally, this phylogeny demands fundamental reconsideration of proposals that relate ratite evolution to continental drift.
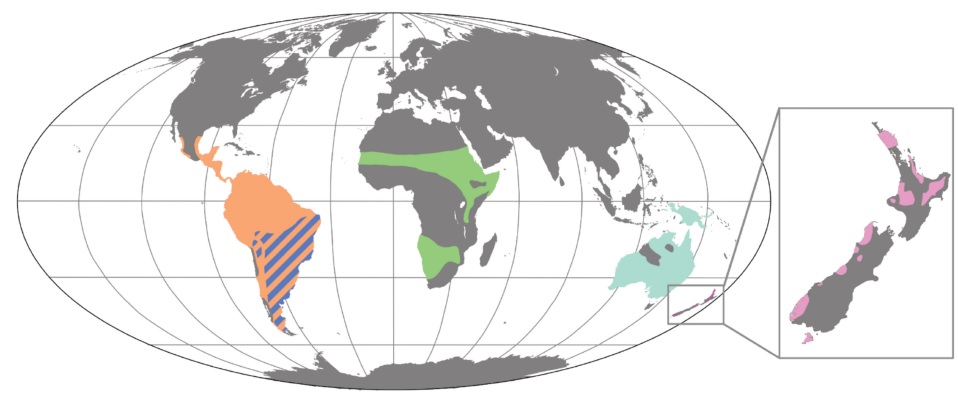
Present-day distribution of extant paleognaths. Dark blue = Rheidae (rheas), orange = Tinamidae (tinamous), green = Struthionidae (Common Ostrich), aqua = Casuariidae (cassowaries),
and pink = Apterygidae (kiwis) (Figure from Widrig and Field 2022).
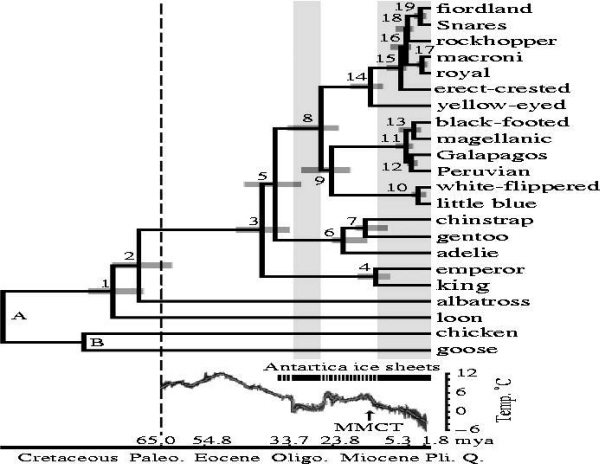
Chronogram of penguin diversification. Nodes A and B indicate 104 and 90 myr. Credibility intervals (95%) are indicated by grey bars at numbered internal nodes. Vertical dashed line indicates the K/T boundary. Periods when Antarctica was ice-covered (black continuous bars) are indicated by shaded grey rectangles. Ocean temperature is based on high-resolution deep-sea oxygen isotope records. The MMCT is indicated by an arrow.
Origin and expansion of penguins -- Classic problems in historical biogeography are where did penguins originate, and why are such mobile birds restricted to the Southern Hemisphere? Competing hypotheses posit they arose in tropical-warm temperate waters, species-diverse cool temperate regions, or in Gondwanaland 100mya when it was further north. To test these hypotheses, Baker et al. (2006) constructed a phylogeny of extant penguins from 5851bp of mitochondrial and nuclear DNA. Their analysis suggests that an Antarctic origin of extant taxa is highly likely, and that more derived taxa occur in lower latitudes. Molecular dating indicates that penguins originated about 71 million years ago (i.e., penguins and albatrosses shared a common ancestor approximately 71 mya) in Gondwanaland when it was further south and cooler (and Antarctica was still attached to Australia and South America, and New Zealand was still relatively close to the supercontinent). Moreover, extant taxa are inferred to have originated about 40 mya (Eocene) when Aptenodytes diverged as the basal lineage, and coincident with the extinction of the larger-bodied fossil taxa as global climate cooled. Baker et al. (2006) hypothesized that, as Antarctica became ice-encrusted, modern penguins expanded via the circumpolar current to oceanic islands within the Antarctic Convergence, and later to the southern continents. Thus, global cooling has had a major impact on penguin evolution, as it has on vertebrates generally. Penguins only reached cooler tropical waters in the Galapagos about 4 mya, and have not crossed the equatorial thermal barrier.
The demise of the larger-bodied putative stem-group taxa near the end of the Eocene about 40 mya coincides roughly with the origin of Antarctic-breeding extant taxa (Aptenodytes and Pygoscelis) in the crown-group, and with the beginning of a general cooling in global climate. This was also approximately when the fish-eating cetaceans evolved, and it has been hypothesized they may have out-competed these larger penguins which probably relied on the same food source. Two abrupt cooling periods resulting in the formation of large ice sheets in Antarctica are associated with the diversification of penguin taxa. The first cooling occurred about 34-25 mya, when Spheniscus, Eudyptes and Eudyptula diverged from the older Antarctic genera. These latter ancestral lineages may have dispersed northward by the newly formed circumpolar current, judging from the occurrence of a Eudyptula fossil in New Zealand about 24 mya. As surface waters in the Southern Oceans continued to cool towards the middle Miocene, and the flow of the circumpolar current around Antarctica intensified, another rapid climate transition, the middle miocene climate transition (MMCT) and subsequent increase in Antarctic ice volume occurred between 14 and 12 mya. The MMCT was accompanied by a second bout of cladogenesis that gave rise to multiple species of extant penguins distributed at even lower latitudes, including tips of southern continents. If this scenario is run backwards in the future, continued global warming might be expected to drive temperate-adapted species out of lower latitudes towards their ancestral distribution, possibly causing multiple extinctions of existing species.
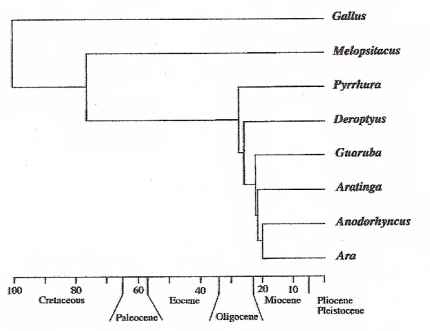
Parrots (Psittaciformes), and other bird orders (e.g., Galliformes; represented by Gallus above), likely diverged about 100 million years ago.
South American parrots (e,g., Pyrrhura, Derptyus, Guaruba, Aratinga, Anodorhyncus, and Ara) diverged from Australian parrots (e.g., Melopsitacus)
about 76 million years ago (Cretaceous-Tertiary boundary) when South America and Australia were moving away from Antarctica (Gondwanaland fission)
(From: Miyaki et al. 1998).
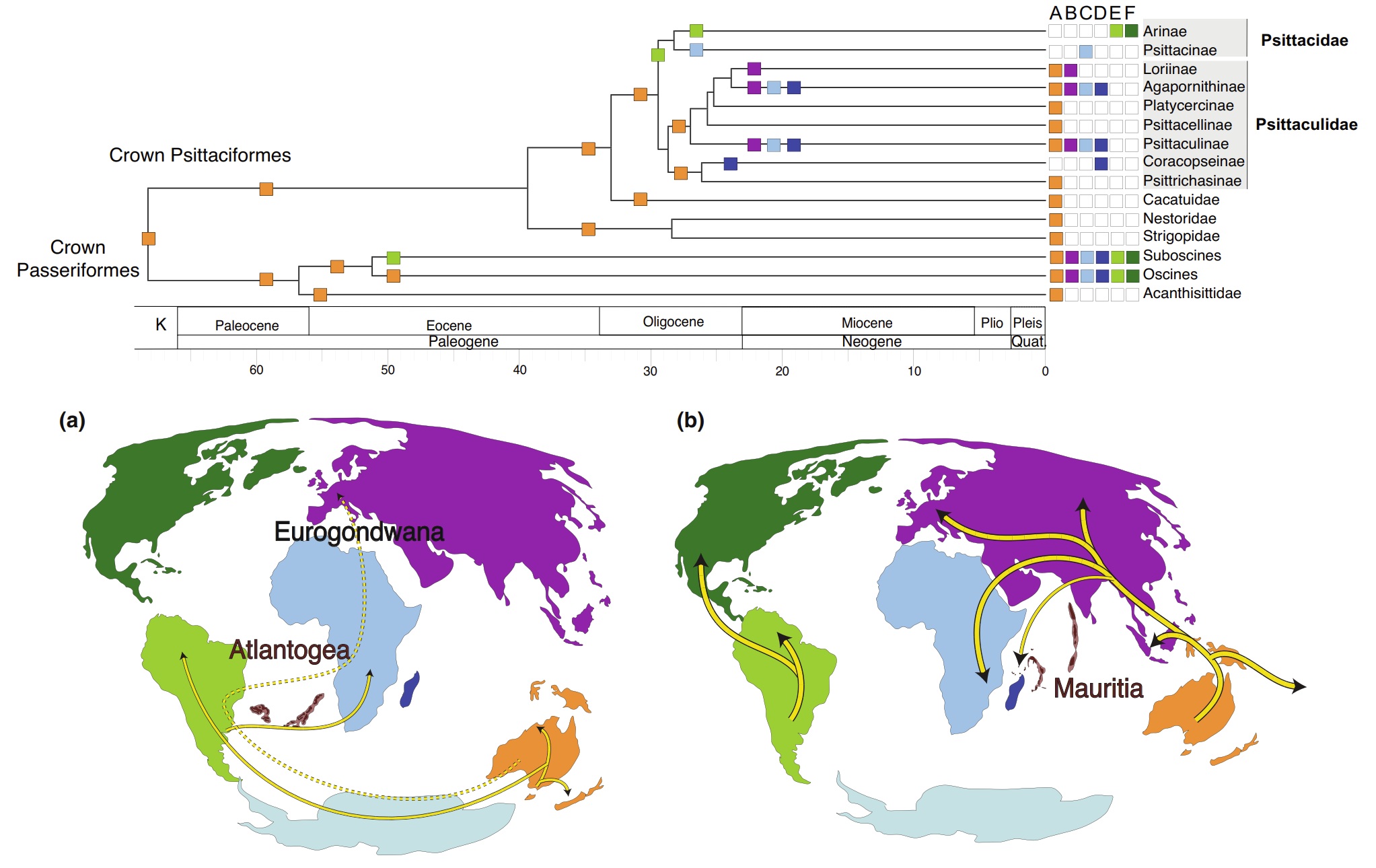
Estimated location of the major landmasses during the (a) Paleogene and (b) Neogene. Yellow lines suggest range expansions/dispersals.
Selvatti et al. (2022) suggested that the largely pantropical distribution of parrots was shaped by two major geological events. First, the
final breakup of parts of Gondwana (separating Australia and South America from Antarctica) may have caused the first splits of parrots,
establishing two parallel radiations: Psittacidae in the Neotropics and Psittaculidae in Australasia. Second, igneous palaeoprovinces (particularly,
Atlantogea in the South Atlantic that connected the Neotropics and Afrotropics) could have connected, and facilitated dispersal between,
South America, Africa and Europe. Colors of landmasses correspond to the locations of taxa in the above phylogeny.
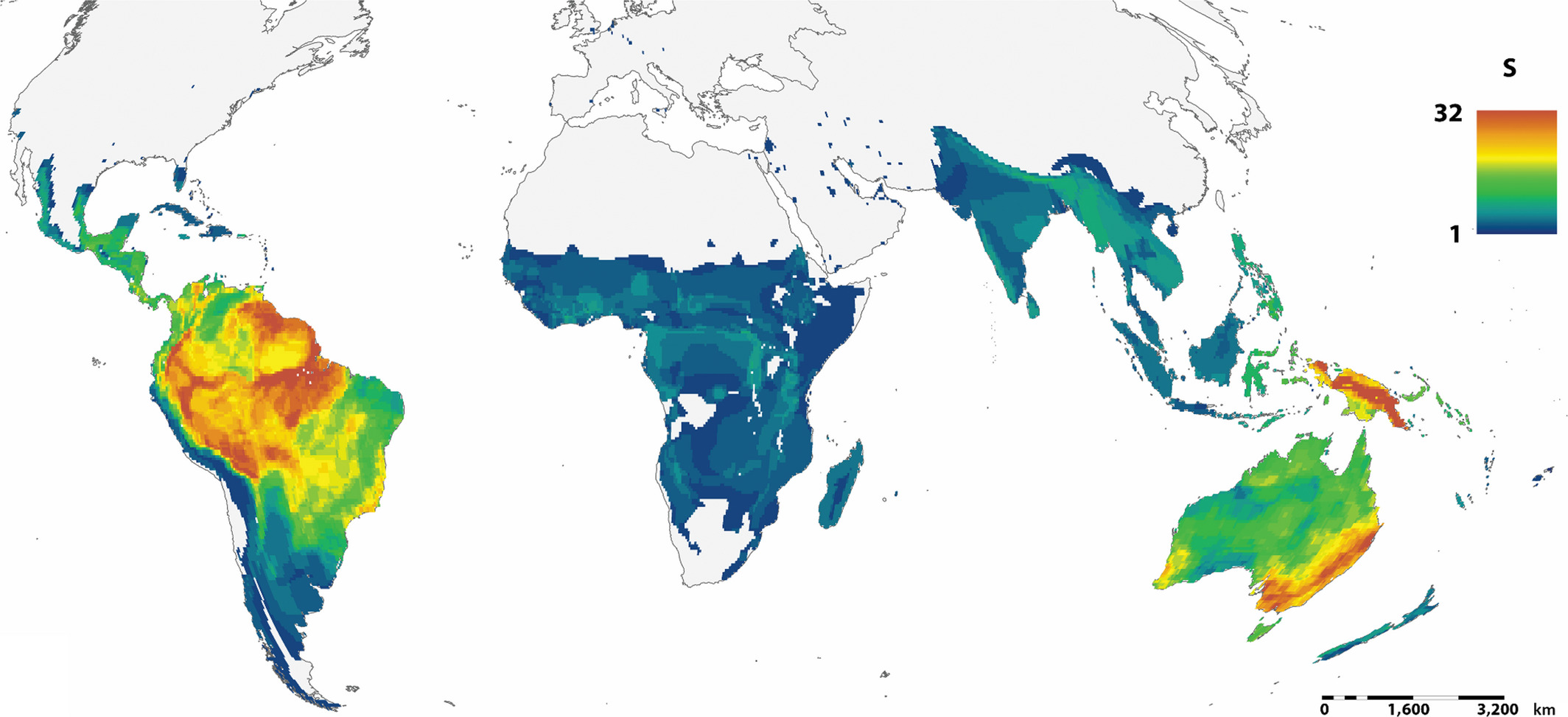
Global variation in parrot species richness. Parrots are distributed circumtropically, with the greatest species
richness in southeastern Australia, Papua New Guinea, Amazonia and northeastern South America (Kosman et al. 2019).
The distribution of a species is a consequence of its:
Given these various factors, how are the world's birds distributed across the six primary zoogeographical
realms, the Nearctic, Neotropical, Palearctic, Ethiopian (or Afrotropical),
Oriental (or Indomalayan),
and Australasian?
| Evaluating species limits in birds -- Watson (2005) examined sources of data used to diagnose new species and found variation among classes. New species of amphibians were typically diagnosed using external appearance, morphometrics, and external morphology, while reptiles were typically diagnosed using appearance and external morphology. Mammals were generally described using morphometrics, external morphology, and internal morphology, whereas new bird species diagnoses relied on external appearance, morphometrics, and behavior (see figure to the right). Birds were the only group described primarily using external appearance (98%; only one species description did not use plumage characteristics), compared with an overall frequency of 54% for the other three groups combined. Ecological data (distributional range, habitat, and behavior) were used for 62% of new bird descriptions, significantly more than the 14% for the other three classes combined. The use of molecular data was consistent across all four classes; by contrast, behavioral traits (typically vocalizations) were used for amphibians and birds but not for reptiles or mammals. |
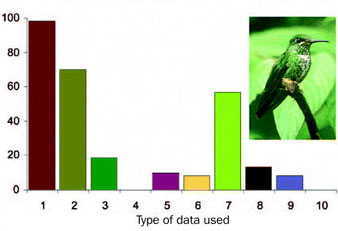 Sources of data used to diagnose new species of birds (1993 - 2002). Data types: 1, appearance; 2, morphometrics; 3, external morphology; 4, internal morphology; 5, range; 6, habitat; 7, behavior; 8, molecular data; 9, karyology; 10, reproduction. Photograph of the Green-crowned Brilliant (Heliodaxa jacula) by David M. Watson. Of the 60 new bird species described, there were 7 owls, 7 tyrant flycatchers, 2 hummingbirds, and 1 parrot (see www.csu.edu.au/faculty/sciagr/eis/staff/watson.htm for details). |

DNA barcode (base pair sequence) of a Green Heron
(Photo courtesy Biodiversity Institute of Ontario)
Birds and DNA barcodes -- The DNA barcodes (see below for an explanation of DNA barcoding) of 643 North American bird species from the Arctic tundra to the temperate woodlands to the Florida Keys (93% of 690 known breeding species in the U.S. and Canada) have now been evaluated (Kerr at al. 2007). According to Paul Hebert of the Biodiversity Institute of Ontario, analysis has revealed that "there are a number of cases of deep genetic divergences within what are currently called single species." These "cryptic species" are those with unique DNA barcodes but differences of song and plumage so subtle as to make them virtually indistinguishable from some other species. Even though birds may appear very similar to human observers, species with distinct DNA barcodes rarely interbreed. "Did we find concordance between barcode results and conventional taxonomy? The answer is, resoundingly, yes. In 95% of cases, entities recognized as species are barcode distinct," adds Dr. Hebert. Dr. Stoeckle of Rockefeller University says there are about 10,000 recognized bird species today and predicts that, at a global scale, DNA barcoding will distinguish at least 1,000 potential new species. The researchers hope to complete an all-bird DNA inventory by 2011. Given the continent's legions of bird specialists, he says he was surprised by the extent of "hidden diversity" revealed in North America, and by the clear DNA distinctions between species. Dr. Stoeckle cautions that "some pairs of listed species now shown to be DNA twins may be relatively young species and prove different over time." As well, he notes, there is no universal scientific agreement on what defines a species.
The importance of this work to conservation is particularly critical, adds Dr. Hebert. For example, the Solitary Sandpiper, its habitat increasingly under pressure from land development and climate change, was not known previously to have two forms, yet their DNA reveals two distinct groups split about 2.5 million years ago. "How can you develop strategies to preserve highly different genetic entities if you don't know they're there? Our work is providing the first molecular evidence of some of these splits." Perhaps the biggest surprise is that DNA barcoding works as well as it does, that a relatively short code of genes (base pairs) distinguishes species so clearly and there isn't more blurring between species, says Dr. Stoeckle. Barcoding has prompted speculation and theories about mechanisms that strip variation out of species and keep them sharply distinct -- periodic "selective sweeps" -- and the reasons behind them. "This work is raising questions about how evolution works and what species are," he says. So far, the Barcode of Life Data Systems ( www.barcodinglife.org ) has catalogued more than 25,000 species of all types, and over 200,000 individual records.
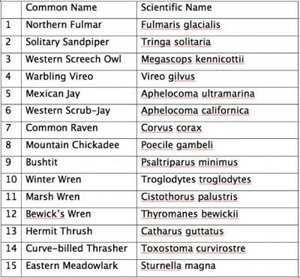
"New" bird species based on DNA barcode analysis (Kerr et al. 2007).
These 15 species each include two distinct barcode clusters.
DNA barcoding is a technique for characterizing species of organisms using a short DNA sequence from a standard and agreed-upon position in the genome. DNA barcode sequences are very short relative to the entire genome and they can be obtained reasonably quickly and cheaply. The cytochrome c oxidase subunit 1 mitochondrial region (COI) is emerging as the standard barcode region for higher animals. It is 648 nucleotide base pairs long in most groups, a very short sequence relative to 3 billion base pairs in the human genome, for example. The ‘barcode’ metaphor is useful though not correct in fine detail. That is, all the products of one type on a supermarket shelf share exactly the same 11-digit barcode that is distinct from all other barcodes. DNA barcodes vary among individuals of the same species, but only to a very minor degree. If the DNA barcode region is effective, the minor variation within species will be much smaller than the differences among species.
Alfred Russell Wallace (1876) studied the world-wide distribution of animals, including birds, and noted that different regions possessed biotas that were, to varying degrees, unique. Based on his analysis, he delineated six such regions, or zoogeographical realms: Nearctic, Palearctic, Neotropical, Ethiopian, Oriental, and Australian. With minor names changes (Figure below), these realms are still recognized as having distinctive assemblages of birds. Each realm has their characteristic birds; avifaunas that represent a mixture of species of various ages & origins. Of course, some birds are found beyond the borders of these realms on Oceanic islands and in Antarctica.
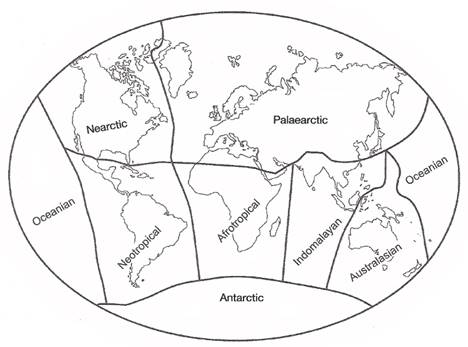
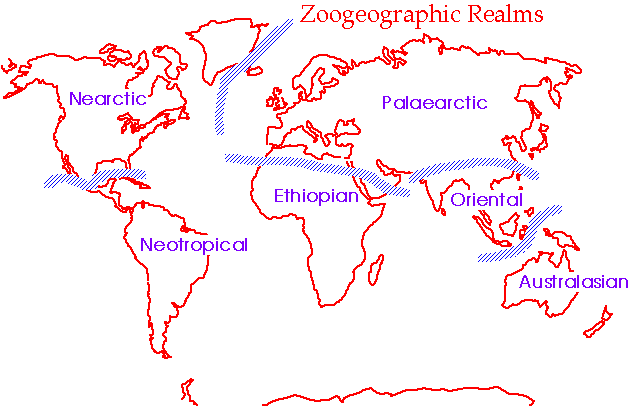
The 'original' zoogeographical realms (Wallace 1876).
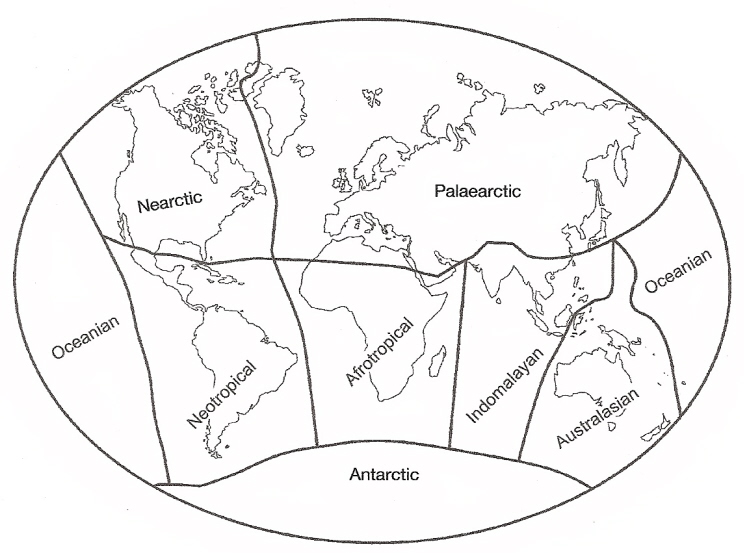
Currently recognized zoogeographical realms and their approximate boundaries (Newton and Dale 2001).
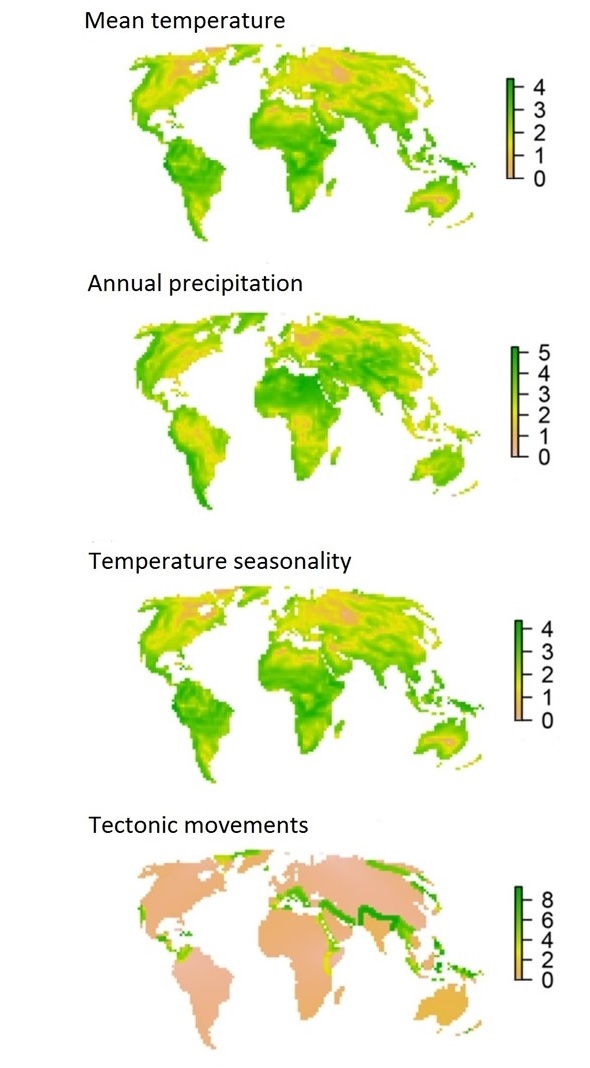
Factors contributing to the location of boundaries between realms
(Figure modified from a supplemental figure from Ficetola et al. 2017).
Here's a brief summary of the avifaunas of these realms:
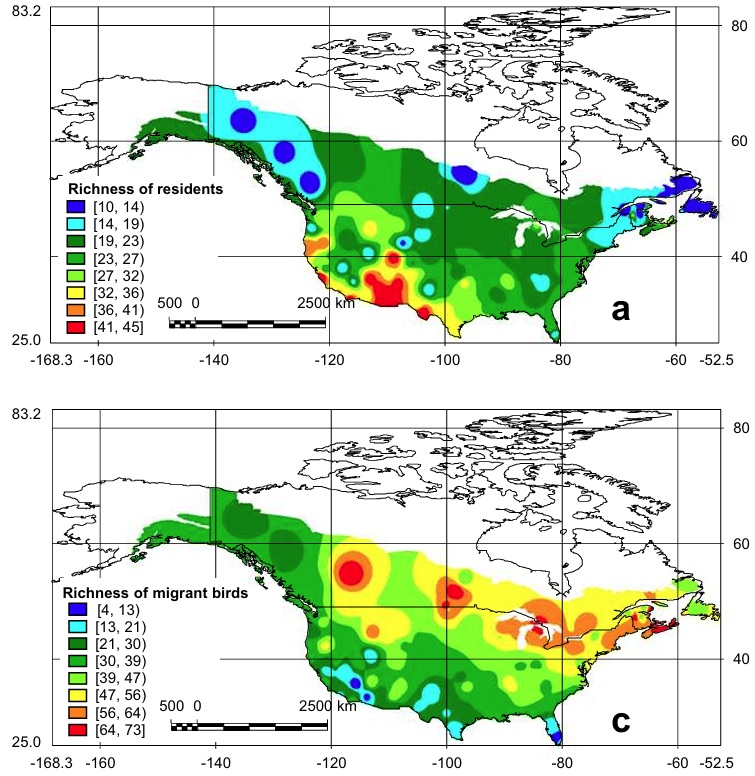
Species richness of resident (top) and migrant (bottom) birds in the United States and Canada. Red color = highest species richness.
Several ecological variables appear to regulate large-scale patterns of terrestrial bird species richness in North America, mainly temperature-, productivity- and landscape habitat structure-related variables. These variables are diverse and group-specific. For instance, the results supported the productivity hypothesis for migratory birds and the winter tolerance hypothesis for resident birds. Habitat structure was also identified as an important factor driving species richness (From: Carnicer and Diaz-Delgado 2008).
| Explosive speciation in Setophaga warblers -- The 27 species of Setophaga wood-warblers represent North America's most spectacular avian adaptive radiation. Setophaga species exhibit high levels of local sympatry and differ in plumage and song, but the group contrasts with other well-known avian adaptive radiations such as the Hawaiian honeycreepers and Galapagos finches in that Setophaga species have differentiated modestly in morphometric traits related to foraging. Instead, sympatric Setophaga tend to partition resources behaviorally and have become a widely cited example of competitive exclusion. Lovette and Bermingham (1999) explored the temporal structure of Setophaga diversification via a phylogeny based on 3639 nucleotides of protein-coding mitochondrial DNA (mtDNA). Comparisons with a null model of random bifurcation-extinction demonstrated that cladogenesis in Setophaga has been clustered non-randomly with respect to time, with a significant burst of speciation occurring early in the history of the genus, possibly as long ago as the Late Miocene or Early Pliocene. In North America, an abrupt rise in temperature & aridity occurred near the Miocene - Pliocene boundary, resulting in a reduction and fragmentation of forest habitats. By the Middle Pliocene, however, the continent-wide distribution of forest habitats was similar to present-day conditions. This sequence of paleoenvironmental changes suggests that the many Setophaga lineages may have initially differentiated allopatrically in the forest refugia of the Early Pliocene, followed by ecological reinforcement of adaptive differentiation during secondary contact in the expanded forests of the Middle Pliocene. | 
|
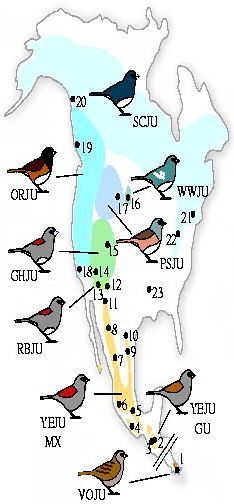 |
Phenotypic and genetic variation in the genus Junco. The eight different taxa included in the genetic analyses are shown. Colors indicate breeding ranges and dots represent sampling localities. From the south, clockwise, with sampling locality number in parentheses: Volcano (1), Yellow-eyed (2-11), Red-backed (12, 13), Grey-headed (14, 15), White-winged (16), Pink-sided (17), Oregon (18, 19) and Slate-colored Juncos (20-23). Site 23 corresponds to a wintering locality of the Slate-colored Junco in Alabama. |
Postglacial range expansion drives the rapid diversification of the genus Junco -- Pleistocene glacial cycles are thought to have played a major role in the diversification of temperate and boreal species of North American birds. Given that coalescence times between sister taxa typically range from 0.1 to 2.0 Myr, it has been assumed that diversification occurred as populations were isolated in refugia over long periods of time, probably spanning one to several full glacial cycles. In contrast, the rapid postglacial range expansions and recolonization of northern latitudes following glacial maxima have received less attention as potential promoters of speciation. Milá et al. (2007) reported a case of extremely rapid diversification in the songbird genus Junco as a result of a single continent-wide range expansion within the last 10000 years. Molecular data from 264 juncos sampled throughout their range reveal that as the Yellow-eyed Junco (Junco phaeonotus) of Mesoamerica expanded northward following the last glacial maximum, it speciated into the Dark-eyed Junco (Junco hyemalis), which subsequently diversified itself into at least five markedly distinct and geographically structured morphotypes in the USA and Canada. Patterns of low genetic structure and diversity in mitochondrial DNA and amplified fragment length polymorphism loci found in Dark-eyed Juncos relative to Mesoamerican Yellow-eyed Juncos provide support for the hypothesis of an expansion from the south, followed by rapid diversification in the north. These results underscore the role of postglacial expansions in promoting diversification and speciation through a mechanism that represents an alternative to traditional modes of Pleistocene speciation. Recent divergence with fast plumage evolution (since the LGM) has been recently documented in Icterus orioles, redpolls, bluethroats, yellow wagtails, and Yellow-rumped Warblers. However, given the higher number and marked distinctiveness of plumage morphs and the short amount of time involved, the Dark-eyed Junco complex represents an exceptional case of fast plumage diversification. Given the marked differences in plumage characters, sexual selection may have acted as the mechanism of divergence.
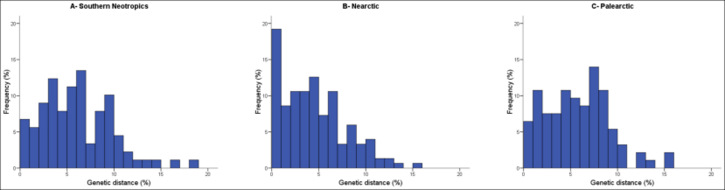
Distribution of nearest congeneric neighbor distances in COI (i.e., cytochrome c oxidase I gene region) for bird species in three biogeographic regions. Distances tend to be higher in the southern Neotropics than in North America and the incidence of species pairs with very low divergence (less than 1% sequence distance) is much higher in North America. Divergence values in the Palearctic are closer to those of the Neotropics than to those in the Nearctic.
Continental patterns of avian diversification -- Causes for the higher biodiversity in the Neotropics as compared to the Nearctic and the factors promoting species diversification in each region have been much debated. The refuge hypothesis posits that high tropical diversity reflects high speciation rates during the Pleistocene, but this conclusion has been challenged. Lijtmaer et al. (2011) investigated this matter by examining continental patterns of avian diversification through the analysis of large-scale DNA barcode libraries. Datasets from the avifaunas of Argentina, the Nearctic, and the Palearctic were analyzed. Average genetic distances between closest congeners and sister species were higher in Argentina than in North America reflecting a much higher percentage of recently diverged species in North America (Nearctic). In the Palearctic, genetic distances between closely related species appeared to be more similar to those of the southern Neotropics. Average intraspecific variation was similar in Argentina and North America, while the Palearctic fauna had a higher value due to a higher percentage of variable species. Geographic patterning of intraspecific structure was more complex in the southern Neotropics than in the Nearctic, while the Palearctic showed an intermediate level of complexity. This analysis suggests that avian species are older in Argentina than in the Nearctic, supporting the idea that the greater diversity of the Neotropical avifauna is not caused by higher recent speciation rates. Species in the Palearctic also appear to be older than those in the Nearctic. These results, combined with the patterns of geographic structuring found in each region, suggest a major impact of Pleistocene glaciations in the Nearctic, a lesser effect in the Palearctic and a mild effect in the southern Neotropics.
In the Nearctic, almost all variable species showed eastern and western lineages hat is consistent with population expansions from glacial refugia in the margins of North America. The eastern and western portions of the Nearctic probably acted as refugia during the early and mid Pleistocene, but were later covered by ice. This is consistent with the fact that the eastern and western lineages within species typically show more than 1.5% divergence. In the Palearctic, three patterns were found and interpreted at least partially as evidence of range expansion from refugia after the retreat of the ice or permafrost that covered much of the northern portion of Eurasia. Finally, in the Neotropics several patterns were found and they appear to reflect various sources of diversification, including the Andes Mountains. These differences once again support the conclusion of a major effect of Pleistocene glaciations in the Nearctic, a less marked and more heterogeneous effect in the Palearctic, and an even milder effect in the southern Neotropics, where other diversification processes may have been as relevant in shaping current diversity.
Rufous-capped Antthrush
To learn more about Neotropical birds (and, more broadly, the birds of the world), visit: Cornell Lab of Ornithology Birds of the World Online!
|
Suboscines and Oscines Of the more than 3,700 species of Neotropical
birds, approximately
1,000 species are classified taxonomically as "suboscines." There are
only
about 50 suboscine species in all of the rest of the world, thus the
Neotropics
are unusual in harboring so many members of this group. The suboscines
are part of the huge order of Passeriformes, or perching birds. Most
passerines
in the world are true oscines, which means that they have a complex
musculature
of the syrinx, the part of the trachea that produces elaborate sounds,
such as the flutelike songs of various thrushes and solitaires or the
warbling
of a canary. Suboscines, however, have a considerably less complex
syringeal
musculature and typically have far more limited singing abilities than
true oscines. Neotropical suboscines have undergone two major adaptive
radiations, with the tyrant flycatchers, cotingas, and manakins
representing
one, and the woodcreepers, ovenbirds, true antbirds, ground antbirds,
gnateaters,
and tapaculos representing the other. No one knows why suboscines have
fared so well in the Neotropics, but the reason may simply be
historical.
For more information about birds of the neotropics, check out John
Kricher: A Neotropical Companion: Chapter 12. Neotropical Birds.
|
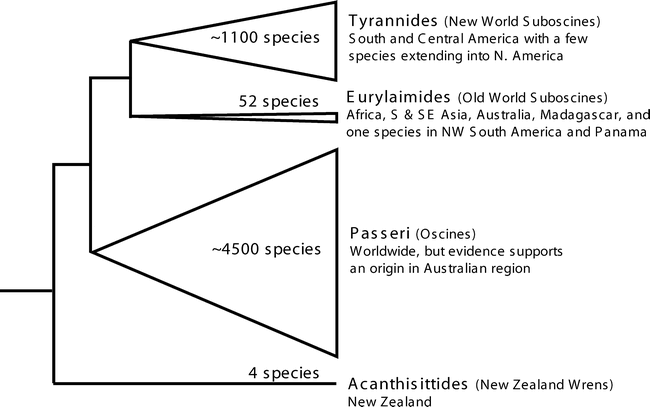
Summary of passerine bird distribution and basal relationships (Moyle et al. 2006)
Chesser (2004) studied the phylogenetic relationships among New World suboscine birds using nuclear and mitochondrial DNA sequences. New World suboscines were shown to constitute two distinct lineages, one apparently consisting of the single species Sapayoa aenigma (Broad-billed Sapayoa), the other made up of the remaining 1000+ species of New World suboscines. With the exception of Sapayoa, monophyly of New World suboscines was strongly corroborated.

Broad-billed Sapayoa
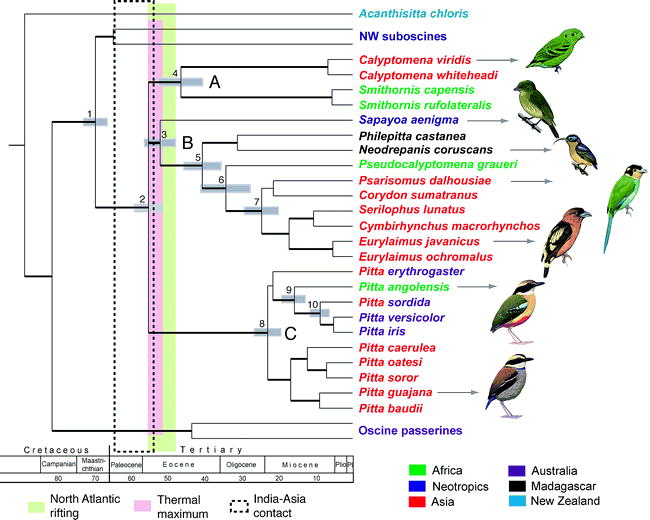
Moyle et al. (2006) used molecular and morphological data to derive a phylogenetic hypothesis for the Eurylaimides, an Old World bird group now known to be distributed pantropically, and to investigate the evolution and biogeography of the group. Phylogenetic results indicated that the Eurylaimides consist of two monophyletic groups, the pittas (Pittidae) and the broadbills (Eurylaimidae sensu lato), and that the broadbills consist of two highly divergent clades, one containing the sister genera Smithornis and Calyptomena, the other containing Pseudocalyptomena graueri, Sapayoa aenigma, the asity genera Philepitta and Neodrepanis, and five Asian genera. Their results indicate that over a ~10 million year time span in the early Tertiary, the Eurylaimides came to inhabit widely disjunct tropical regions and evolved disparate morphology, diet, and breeding behavior. Biogeographically, although a southern origin for the lineage is likely, time estimates for major lineage splitting do not correspond to Gondwanan vicariance events, and the biogeographic history of the crown clade is better explained by Laurasian climatic and geological processes. In particular, the timing and phylogenetic pattern suggest a likely Laurasian origin for the sole New World representative of the group, Sapayoa aenigma.
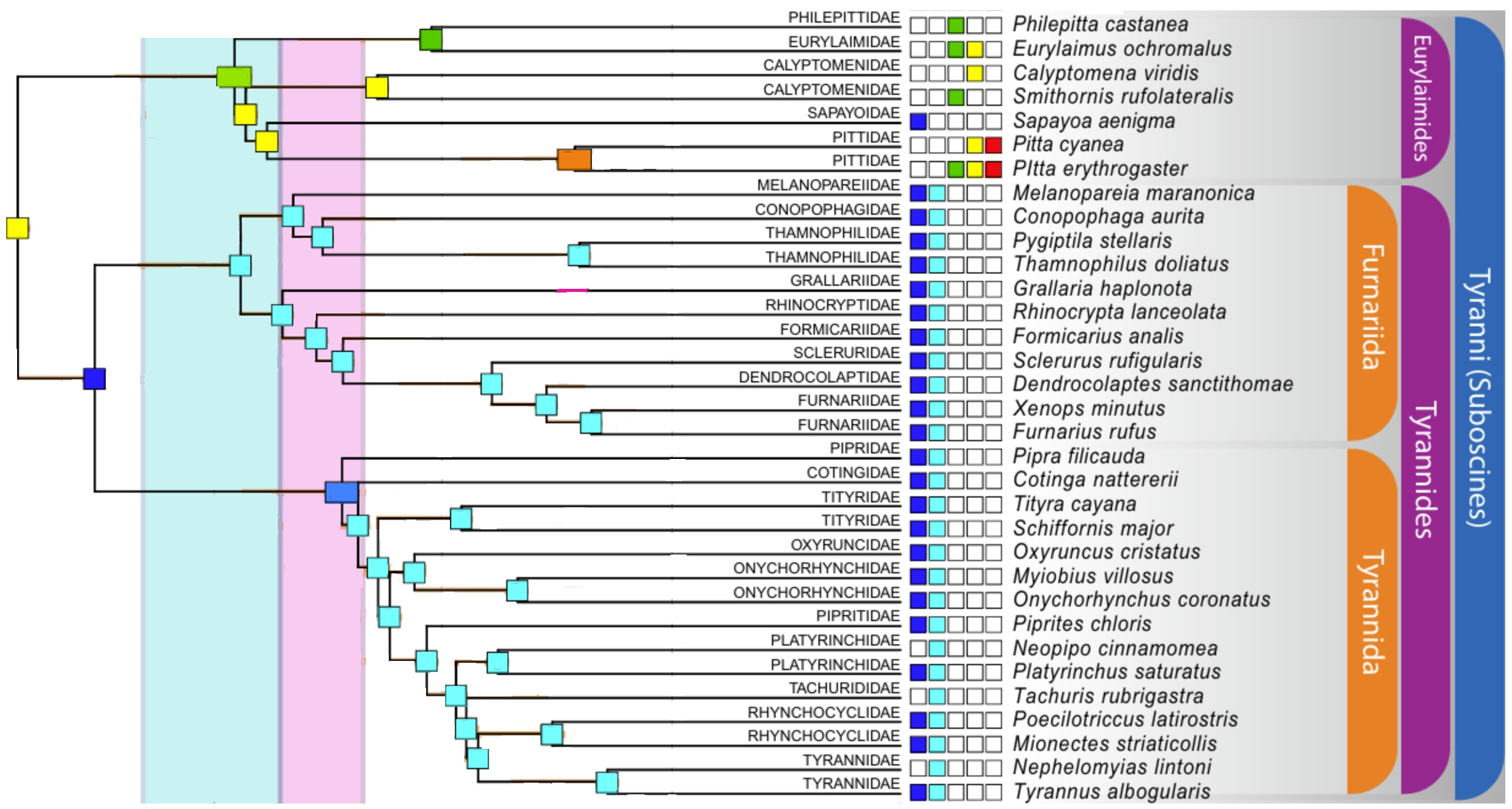
Family-level phylogenetic relationships of suboscine passerines. Colored squares to the left of each species indicate
the continent (see Figure above the phylogeny) where the species is found. Eurylaimides = Old World suboscines; Tyrannides =
New World suboscines. Note that Sapayoa aenigma is the only Old World suboscine found in the New World (From: Oliveros et al. 2019).
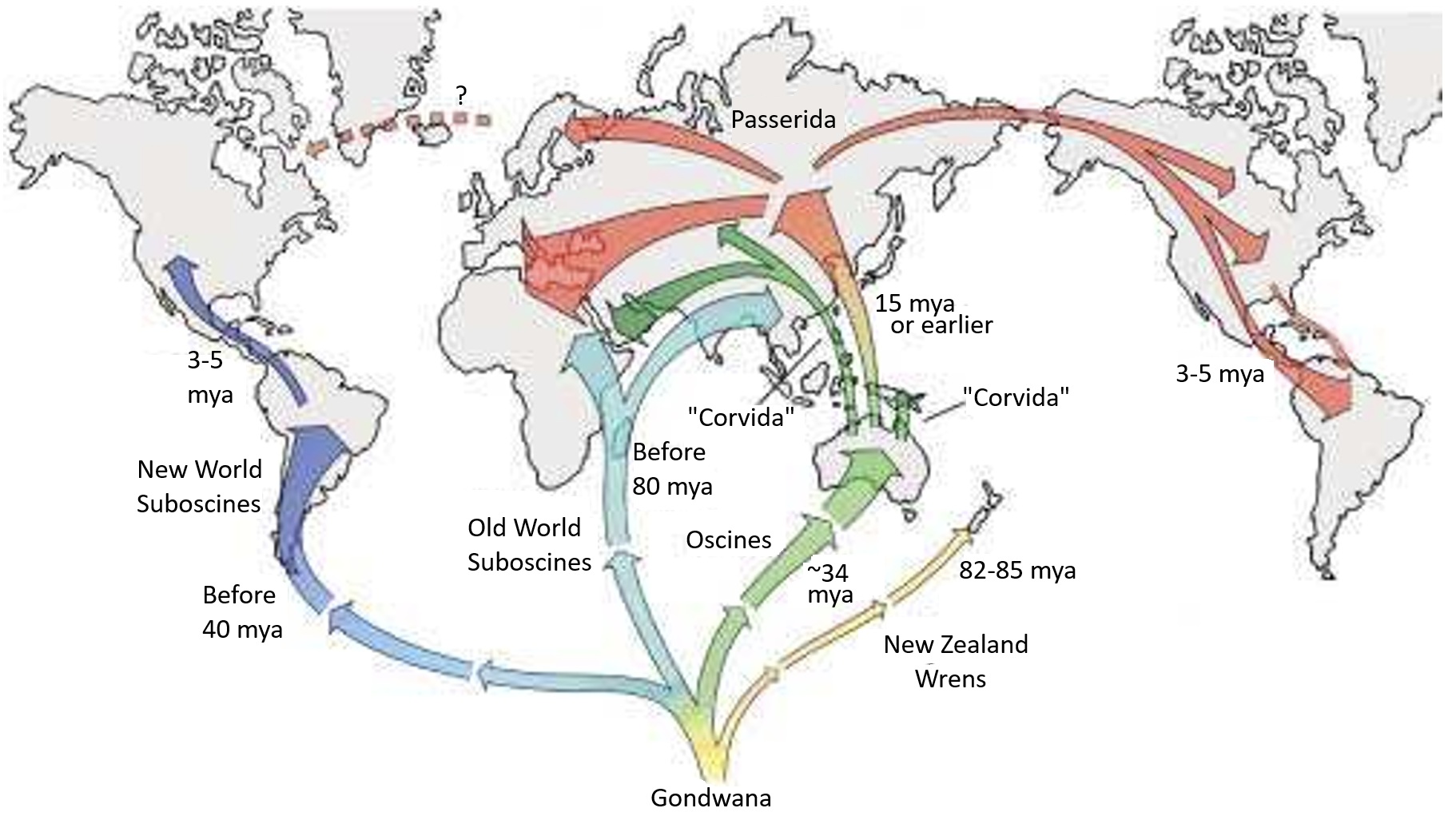
Possible dispersal routes of major passerine groups from Gondwana based on phylogenetic and biogeographic data.
The southern continents are shown in their current position, with Gondwana as a central southern origination point
for clarity. Note the possible dispersal routes of Old World and New World suboscines (From: Ericson et al. 2003).
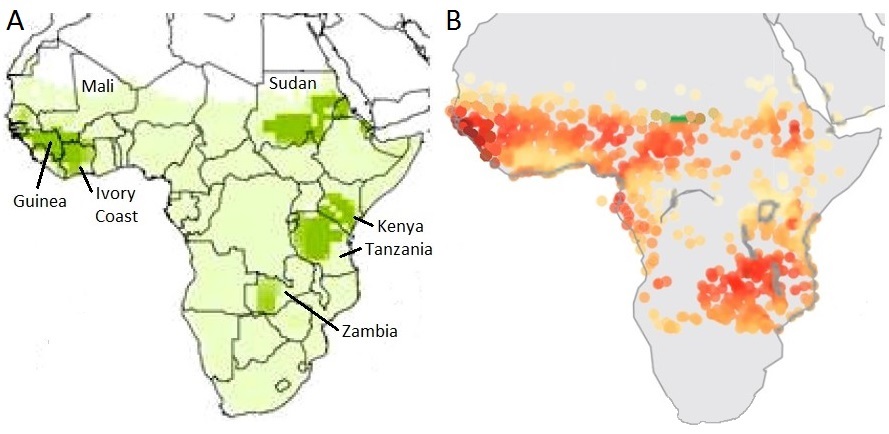
(A) Areas where 68 species of Palearctic migrant songbirds winter in sub-Sahara Africa, with darker green indicating a greater number of species.
(B) Areas with the most seasonal patterns of rainfall in sub-Sahara Africa (with darker red indicating greater seasonality), with rainfall peaking at the beginning of the
wintering period for Palearctic migrants (Figure A modified fom Walther and Pirsig 2017 and B modified from Feng et al. 2013).
Understanding migratory connectivity of Palearctic-African birds and the conservation consequences
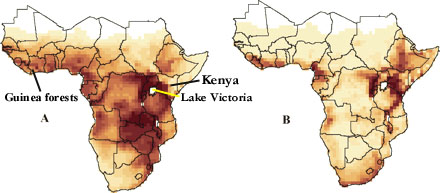
Species richness of breeding birds endemic to Africa
for species with large
(>100 1o quadrats,
840 species, A) and small (100 1o
quadrats, 756 species, B) range size.
Values range from 2 (light) to 440 (dark) in A and from
0 (light) to 120 (dark) in B
(From: Jetz and Rahbek 2001).
Although at a similar latitude than the Neotropical realm, the Afrotropical realm has fewer than half as many species and fewer endemics. Among the taxa thought to have arisen in Africa are Struthionidae (Ostrich), Sagittariidae (Secretarybird, Sagittarius serpentarius), Scopidae (Hammerkop), Balaenicipitidae (Shoebill), Musophagidae (turacos), Coliidae (mousebirds), Phoeniculidae (wood-hoopoes), scimitarbills (Rhinopomastidae), Numididae (guineafowl), Prionopidae (helmet-shrikes), honeyguides (Indicatoridae), and most bee-eaters (Meropidae). The Old World suboscines (the genera Smithornis and Pitta plus Grauer’s Broadbill, Pseudocalyptomena graueri) may also have originated in Africa (Cracraft 1973, Fjeldså and Bowie 2008). Most of these endemic taxa include fewer than 10 species (Cracraft 1973).
Madagascar is also part of the Afrotropical realm. The current avifauna of Madagascar consists of 258 species (with 209 of those species breeding in Madagascar which, compared to land masses of similar size, is rather species poor (Reddy et al. 2012). However, because nearly half of the species in Madagascar are endemic (109 species) and are found nowhere else (McDowell 2008), the avifauna of Madagascar is unique and of important conservation concern. The only oceanic island comparable to Madagascar in terms of the percentage of endemic species of birds is New Zealand (40% endemism, 8 endemic families; McDowall 2008). Most species of birds in Madagascar, including 80 of the 109 endemic species, are found only in forest habitats (Morris and Hawkins 1998). Since the arrival of humans on Madagascar about 2500 years ago (Crowley 2010), at least 21 endemic species have become extinct (McDowall 2008), including the largest ratite species, the Elephant Bird (Aepyornis maximus; 2-3 m tall and up to 275 kg; Hume and Walters 2012) and 11 of other species in the same genus (Dorst 1972). Three of the world's most endangered or vulnerable birds of prey are found in Madagascar, including the Madagascar Serpent-Eagle (Eutriorchis astur), Madagascar Red Owl (Tyto soumagnei), and Madagascar Fish-Eagle (Haliaeetus vociferoides).
Extinct Elephant Bird of Madagascar
Madagascar is rather species poor because it has been isolated since about 84 million years ago when it split from the India landmass. Although its present position is much closer to Africa (about 400 km) than India (4000 km), Madagascar has been separated from Africa since about 150 million years ago (Hallam 1994). The ancestors of most Madagascar birds were from either the IndoMalaya region or, more often, Africa and arrived within the past five million years (Fuchs et al. 2013). However, some colonization appears to have occurred much earlier, including the ancestors of ground-rollers (Brachypteraciidae) sometime between 85 to 49 million years ago (Brown et al. 2008), mesites (Mesitornithidae) about 55 million years ago (Ericson et al. 2006), asities (Eurylaimidae) about 42 million years ago (Proches and Ramdhani 2012), Malagasy warblers (Bernieridae) about 25 million years ago (Proches and Ramdhani 2012), and vangas (Vangidae) about 20 million years ago (Proches and Ramdhani 2012).
Most non-endemic species of Malagasy birds are also shared with Africa, including Marsh Owls (Asio capensis), Madagascar Bee-eaters (Merops superciliosus), and Wattled Starlings (Creatophora cinerea). However, the ancestors of a few species, including the Madagascar Paradise-flycatcher (Terpsiphone mutate), Madagascar Bulbul (Hypsipetes madagascariensis), and Madagascar Magpie-Robin (Copsychus albospecularis), originated in southeast Asia (Marks and Willard 2005).
The birds of Madagascar
Although relatively species poor, some taxa have radiated to exploit some of the diverse niches available in Madagascar. For example, the endemic Malagasy couas (Coua spp.) likely arrived in Madagascar sometime during the Miocene (between 23 and 5 million years ago) and adaptively radiated into 10 species, including nine extant species, most of which have adapted to a specific type of Malagasy forest (Erritzøe et al. 2012). The ancestors of the 11 species in the family Bernieridae colonized Madagascar about 25 million years ago and subsequently diversified. All species in the family glean for insects, but differ in habitat use, feeding strata, morphology, and diet that indicates adaptation to different niches (Hawkins 1999, Schulenberg 2003).
The vangas (Vangidae) represent the largest taxa of endemic songbirds in Madagascar, with 15 genera and 22 species. Most vangas are insectivorous, but some eat fruits and even small vertebrates (Schulenberg 2003). Vangas exhibit impressive diversity in bill morphology that corresponds to difference in foraging behavior and diet (Figure below). Some species probe for insects in bark like woodpeckers and nuthatches (Schulenberg 2003).Moreau (1966) hypothesized that the absence of woodpeckers in Madagascar allowed vangas to radiate into the bark-probing niche.
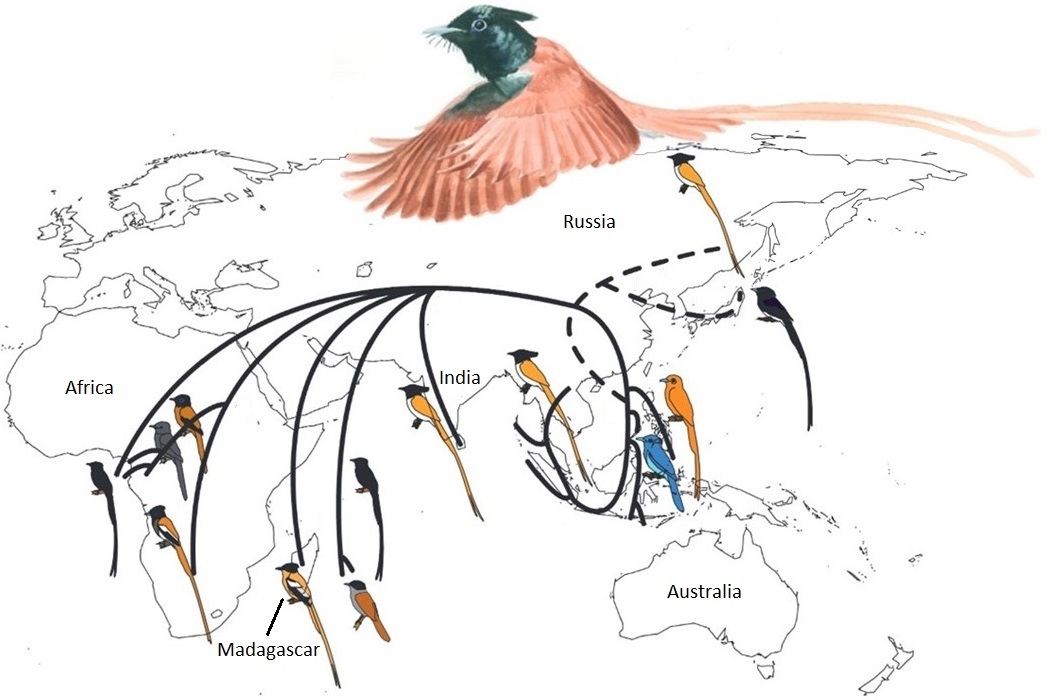
The ancestors of some species of birds in Madagascar originated in southeast Asia. One example is the Madagascar Paradise-flycatcher (Terpsiphone mutate).
Illustrated here are the inferred dispersal and colonization routes of the Terspsiphone lineages. The ancestor of the Madagascar Paradise-flycatcher is
thought to have colonized Madagascar about 2 million years ago. The dispersal and colonization routes of paradise-flycatchers in eastern Asia
are less clear (as indicated by dashed lines) (Figure made from two figures and modified from Fabre et al. 2012).
Oriental (or Indomalayan)
India, southeast Asia (Malay Archipelago), Java, Borneo, the Phillipines, and western Indonesia
~ 2400 species of birds
of 66 families of land & fresh-water birds resident in this region, only one, the Irenidae (Leafbirds & Fairy Bluebirds), is endemic
characteristic birds include many pheasants & pigeons, owls, woodpeckers, pittas, hoopoes, barbets, honeyguides, & sunbirds.
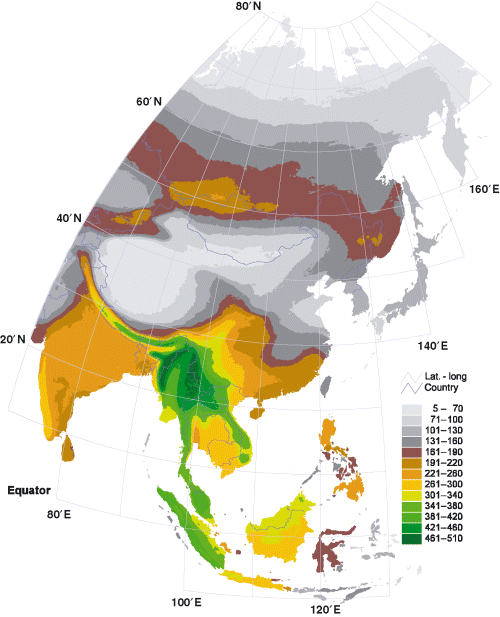
Avian species richness in the Indomalayan realm -- Species richness map overlaid by the breeding ranges of 2406 bird species in east Asia. Bird species richness was highest at the base of the Indochina Peninsula and lowest in northern Siberia, the Gobi and Taklimakan deserts and the Tibetan Plateau. Islands had lower bird species richness than adjacent areas in mainland Asia (Ding et al. 2006).
Ding et al. (2006) suggested that primary productivity is the key factor underpinning patterns of bird species richness in east Asia. Primary productivity sets the upper limits of the capacity of habitats to support bird species. In isolated areas such as islands and peninsulas, however, BSR might not reach the richness limits set by primary productivity because the degree of isolation and area size also can affect species richness. Other factors, such as spatial heterogeneity, biotic interactions, and perturbations, may also affect species richness. However, their effects are secondary and are not as strong as primary productivity, isolation, and area size.
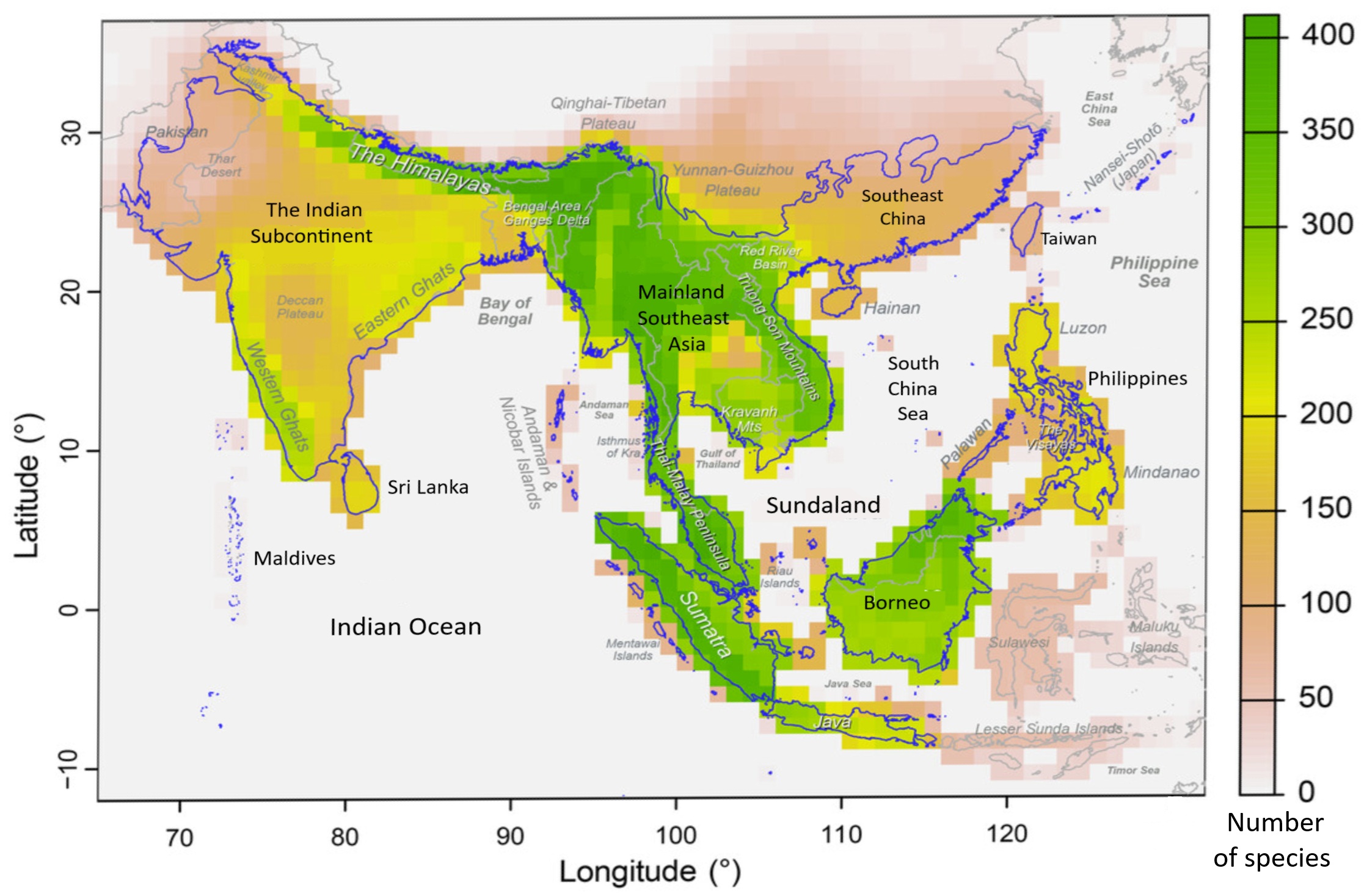
Map showing Indomalaya avian species richness for native resident species (n = 1544).
Oong et al. (2026) examined the biogeographical patterns of birds within Indomalaya. Their analyses revealed four major biogeographic regions in Indomalaya that were consistent with present knowledge about the geological history of the region. (1) Indian Subcontinent. Analysis consistently indicated that the Indian subcontinent is a distinct biogeographic region in Indomalaya, with evidence of local areas of endemism. For example, Sri Lanka and the Western Ghats have both been noted as regions of high endemism34 of the 479 bird species on Sri Lanka are endemic to the island, and the Western Ghats has about 20 endemics. Analysis also revealed evidence of habitat-based subdivision in the Indian subcontinent. The central and western portion of the Indian peninsula is more arid and highly seasonal, whereas the southern and eastern portions around the Western and Eastern Ghats are wetter and relatively less seasonal. (2) Continental Southeast Asia (mainland Southeast Asia and southeastern China). Mainland Southeast Asiaincluding Vietnam, Laos, Cambodia, Thailand, and Myanmar- extends west into the Himalayas. At least 200 bird species have overlapping ranges in this region. Southeastern China includes a hilly coastal strip south of the Yangtze River basin; approximately 50 bird species have overlapping ranges here. (3) Sundaland (Thai-Malay Peninsula, the islands of Sumatra, Borneo, Java, and Palawan, plus several smaller offshore islands). This region is characterized by extensive tropical rainforests that may have developed by the early Miocene and become largely isolated from the rest of Indomalaya with more seasonal habitats to the north and oceans on all other sides. In addition, the region was later subject to repeated sea level fluctuations during the Plio-Pleistocene, altering land cover and connection among the major landmasses. Sundaland is characterised by a high level of endemism, with over 260 endemic land bird species. (4) Oceanic Philippines. The oceanic Philippines is relatively young and did not come into existence until the Eocene at 35 Ma, while its present-day form did not emerge until around 5 Ma (Hall 2012, 2013; Yumul et al. 2003, 2008, 2009). With its history of geological isolation and position at the crossroads between Australasia and the rest of Indomalaya, the Philippines is faunally distinct, with a high level of both species richness and endemism. About 250 species of birds are either endemic or near endemic to the Philippines.
Some Indomalayan groups, e.g., barbets, range as far east as Bali and no further, whereas the range of several Australian birds (e.g., honey-eaters and cockatoos) range as far west as Lombok (35 km from Bali), but not further. This area of rather abrupt change in faunas was first noticed by Alfred Russel Wallace in the mid-1800’s. He observed an expected decline in the proportion of Asian to Australian birds as he traveled west to east through Indonesia, Malaysia, and Brunei, but noted a more substantial shift in that proportion between the islands of Bali and Lombok (islands just 25 km apart) and between Borneo and Sulawesi (or Celebs). The location of the rather abrupt change in species distributions was subsequently called Wallace’s Line. Wallace concluded that the islands west of the line were once connected to each other and to the Asian mainland and the islands east of the line had been similarly connected, but the two areas had always been separate (Wallace 1859). More recent work suggests that the area between Wallace’s Line and Lydekker’s Line (the western boundary of a strictly Australian fauna), called Wallacea, is best considered an area of interchange between Asiatic and Australian floras and faunas, where organisms exhibit a high degree of endemism (Moss and Wilson 1998).
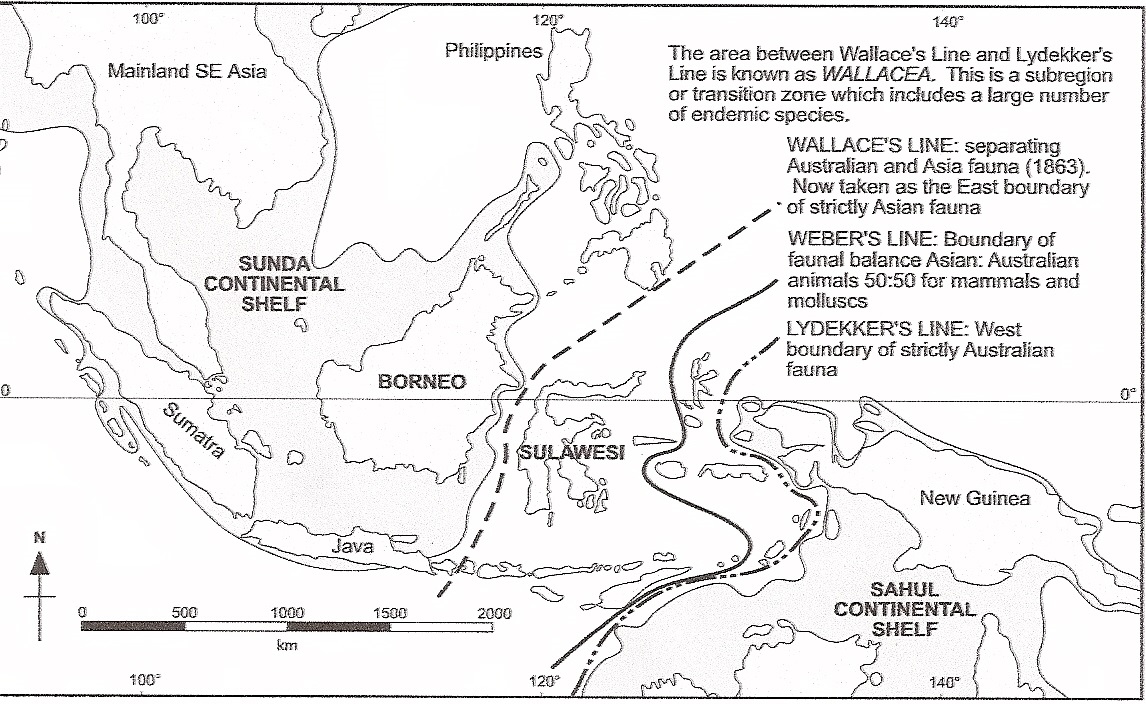
Map of southeast Asia showing the location of Borneo and Sulawesi and Wallace’s Line. East of Java, Wallace’s Line lies between the
islands of Bali and Lombok. Areas of continental shelves are shown in grey (Figure from Moss and Wilson 1998).
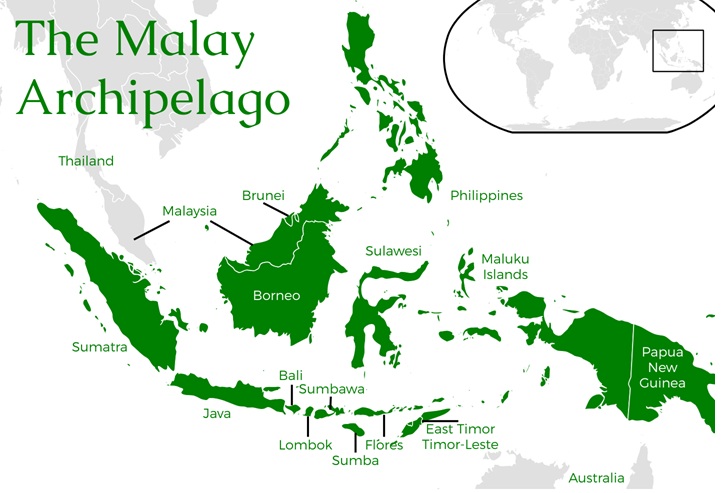
The Malay Archipelago, part of the Sundaland biodiversity hotspot, has exceptionally high bird diversity, hosting over 13% of global bird species. The region, including Sumatra, Java, and Borneo, features rich lowland rainforest assemblages with hundreds of species, high rates of endemismespecially in Borneoand significant populations of hornbills, pittas, and broadbills.
Key Aspects of Bird Diversity in the Malay Archipelago:

Phylogeny of all passerine birds (Order: Passeriformes; n = 1633 species) in the Indo-Australian Archipelago (Figure from: Prasetya 2024).
Iconic and Endemic Species of the Indo-Australian Archipelago:
Key Avian Families and Groups:
Australasian
Australia, New Zealand, Papua New Guinea, eastern Indonesia & several small islands in the area
generally a dry region, only 4.5% rainforest
about 70% of Australia is desert & in this area only 17 species of birds occur (3% of Australia's avifauna)
~ 1600 species in the Australian region (907 in Australia alone, including 642 breeding birds, 162 vagrants, 77 non-breeding birds, and 26 introduced by humans)
~64 families of terrestrial & freshwater birds represented, with several unique:
Cassowaries (Casuariidae) (Learn more about Cassowaries: Part 1, Part 2, Part 3, Part 4).
Emus (Drommiceidae)
Kiwis (Apterygidae)
Owlet-nightjars (Aegothelidae)
New Zealand wrens (Acanthisittidae)
Lyrebirds (Menuridae)
Scrub-birds (Atrichornithidae)
Birds of Paradise (Paradisaeidae)
Bowerbirds (Ptilonorhynchidae)
Honeyeaters (Meliphagidae)
Corcoracidae (two species, the White-winged Chough and Apostlebird)
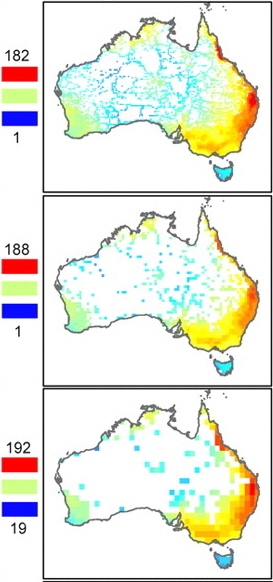 |
Spatial patterns of avian species richness in Australia across three levels of spatial resolution, from 0.25° grid cells to 1° grid cells (Hurlbert and Jetz 2007). The contemporary richness pattern is well explained statistically by actual evapotranspiration (a measure of water–energy balance), operating both directly and indirectly through plant production (Hawkins et al. 2005). |
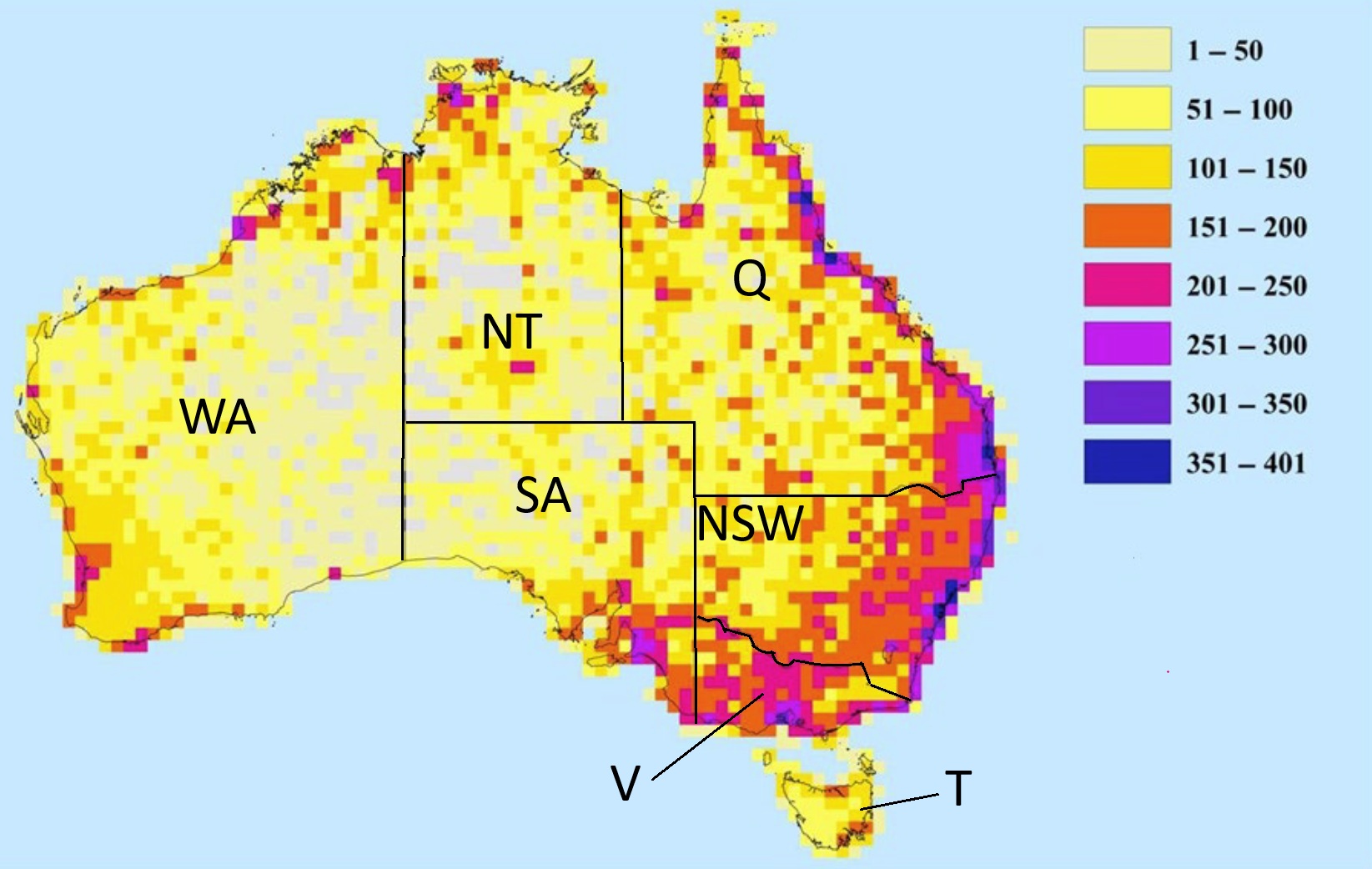
Native bird species richness in Australia at about 50 km x 50 km resolution. Australia has about 830850
bird species, with approximately 45% being endemic, representing nearly one-tenth of the world's total.
Richness is heavily concentrated in the tropical northeast (Queensland) and regions with high, consistent
moisture (high actual evapotranspiration). In addition, structurally complex habitats, such as those with
diverse foliage profiles or larger tree diameters, are strong predictors of higher bird diversity. Within Australia,
Queensland (Q) has the highest bird diversity, with over 630 species recordedroughly 60 more than New South
Wales (NSW), the next most species-rich state Key groups include parrots, honeyeaters, and pigeons. WA. Western
Australia; NT, Northern Territory; SA, South Australia; V, Victoria; T, Tasmania (Figure from: McKinney and Kark 2017).
Australia's Eastern Whipbird
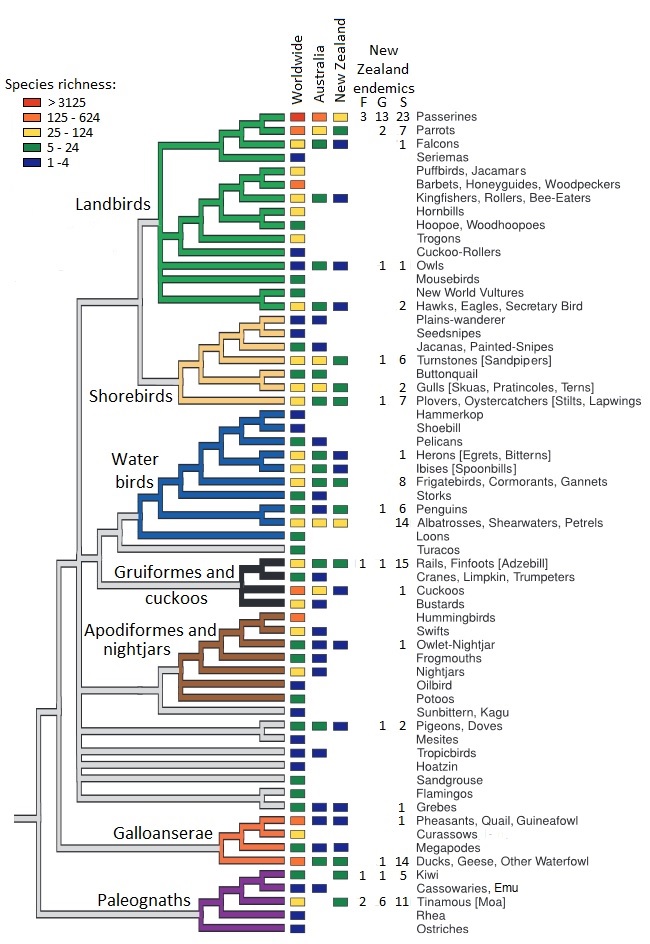
Phylogeny and diversity of birds in Australia and New Zealand, including the number of endemic families (F), genera (G),
and species (S) in New Zealand (Figure modified a bit from Trewick and Gibb 2010).
Before the arrival of humans about 700-800 years ago, there were no terrestrial mammals on New Zealand (and just three species of bats). At that time, the avifauna of New Zealand included at least 245 species and, of these, 59 are now extinct. In the absence of mammals, birds dominated the New Zealand ecosystem. That changed with the arrival of Polynesian Maori settlers. Many species of birds were hunted by these settlers, including moas, shags, parakeets, penguins, pigeons, petrels, ducks, rails, and kiwis (McGlone 1989). As is also the case for many islands in the Pacific, this hunting, along with habitat degradation and introduction of non-native species by the Polynesians that preyed on birds and their eggs led to the extinction of several species. Among the extinct species are the moas, South Island Adzebill (Aptornis defossor), New Zealand Raven ( Corvus antipodum), Waitaha Penguin ( Megadyptes waitaha), Scarlett’s Shearwater (Puffinus spelaeus), Grey-headed Blackbird (Turdus poliocephalus), the world’s largest eagle (Haast’s Eagle, Harpagornis moorei; about 12 kg and a wingspan up to 3 m), and many more.
New Zealand has relatively few native species of landbirds, but does have one endemic order of birds (five species of kiwis, Apterygiformes) two endemic families (two species of New Zealand wrens, Acanthisittidae; two species of New Zealand wattlebirds, Callaeaidae), and 35 endemic species of birds. The ancestors of most species of birds in New Zealand came from Australia (located about 2000 km to the east), aided by the prevailing westerly winds. Evidence of this is provided by several species of birds that have successfully colonized New Zealand from Australia relatively recently, including Silvereyes (Zostertops lateralis, in 1856), Pacific Swallows (Hirundo tahitica, in 1958), Cattle Egrets (Bubulcus ibis, in 1963), Masked Plovers (Vanellus miles, in 1932), Australian Coots (Fulica atra australis, in 1954), White-faced Herons (Ardea novaehollandiae, in 1940) and Black-fronted Dotterels (Charadrius melanops, in 1954) (Falla 1953, Baker 1991). Ancestors of other species of birds in New Zealand arrived from the Holarctic, including the South Island Pied Oystercatcher (Haematopus ostralegus finschi), New Zealand Scaup (Aythya novaeseelandiae), and Auckland Island Merganser (Mergus australis). Surprisingly, the closest ancestor of New Zealand’s only surviving endemic raptor, the Kārearea, or New Zealand Falcon (Falco novaeseelandiae), is the Aplomado Falcon (Falco femoralis) of South America (Trewick and Olley 2016). More recently, at least 36 species of birds have been introduced to New Zealand by humans (Tennyson 2010, Trewick and Gibb 2010).
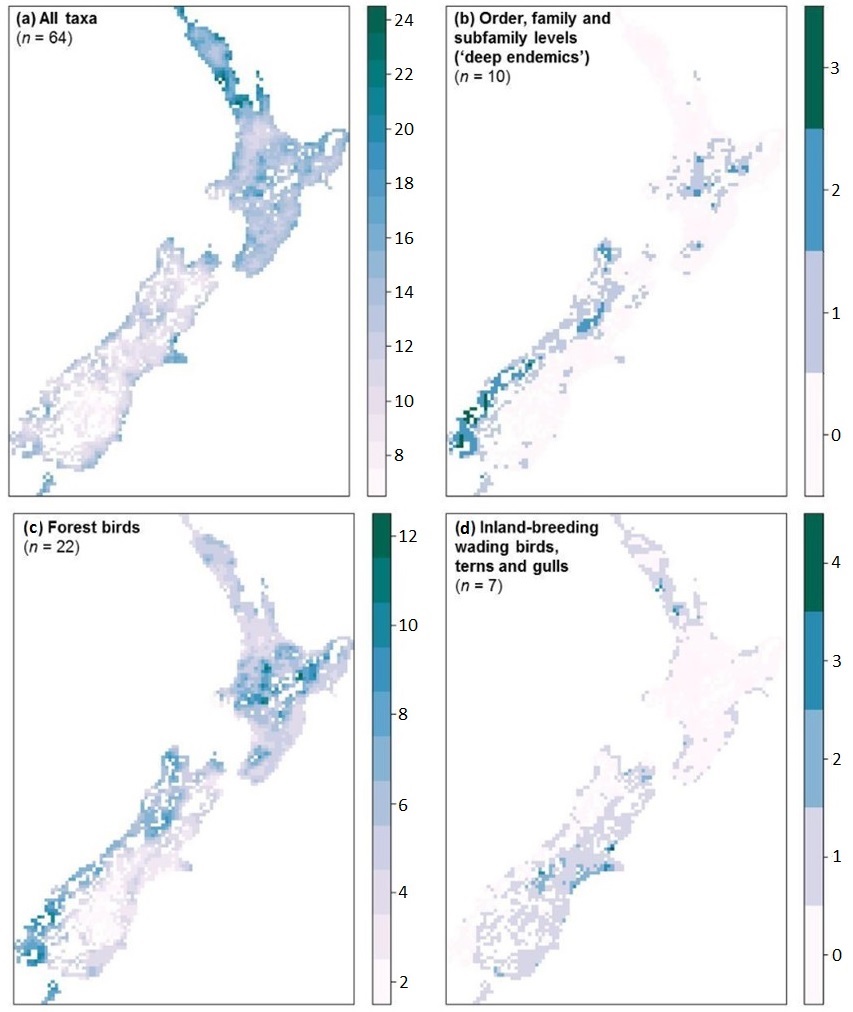
Avian species richness in New Zealand. (a) All native species of land birds, (b) endemic orders, families, and subfamilies, (c) native species of forest birds, and
(d) native species of inland-breeding shorebirds, terns, and gulls. For all native species, species richness is higher further to the north and closer to the coast,
indicating that the diversity of native birds is generally higher where temperatures are higher and where there are coastal-specialist birds in addition to some
land birds. However, species richness of ‘deep-endemic’ species (species in endemic orders, families, and subfamilies), including the
Blue Duck or Whio (Hymenolaimus malacorhynchos), kiwis (Apterygiformes), Kōkako (Callaeas wilsoni), Rifleman (Acanthisitta chloris),
and Rock Wren (Xenicus gilviventris), is higher on New Zealand’s South Island and its smaller nearshore islands, particularly in areas with more forest cover,
less human development, and fewer roads (Figures from Walker and Monks 2017).
Amazing birds found in New Zealand
Evolution and Conservation of Island Birds - Lessons from Genomics
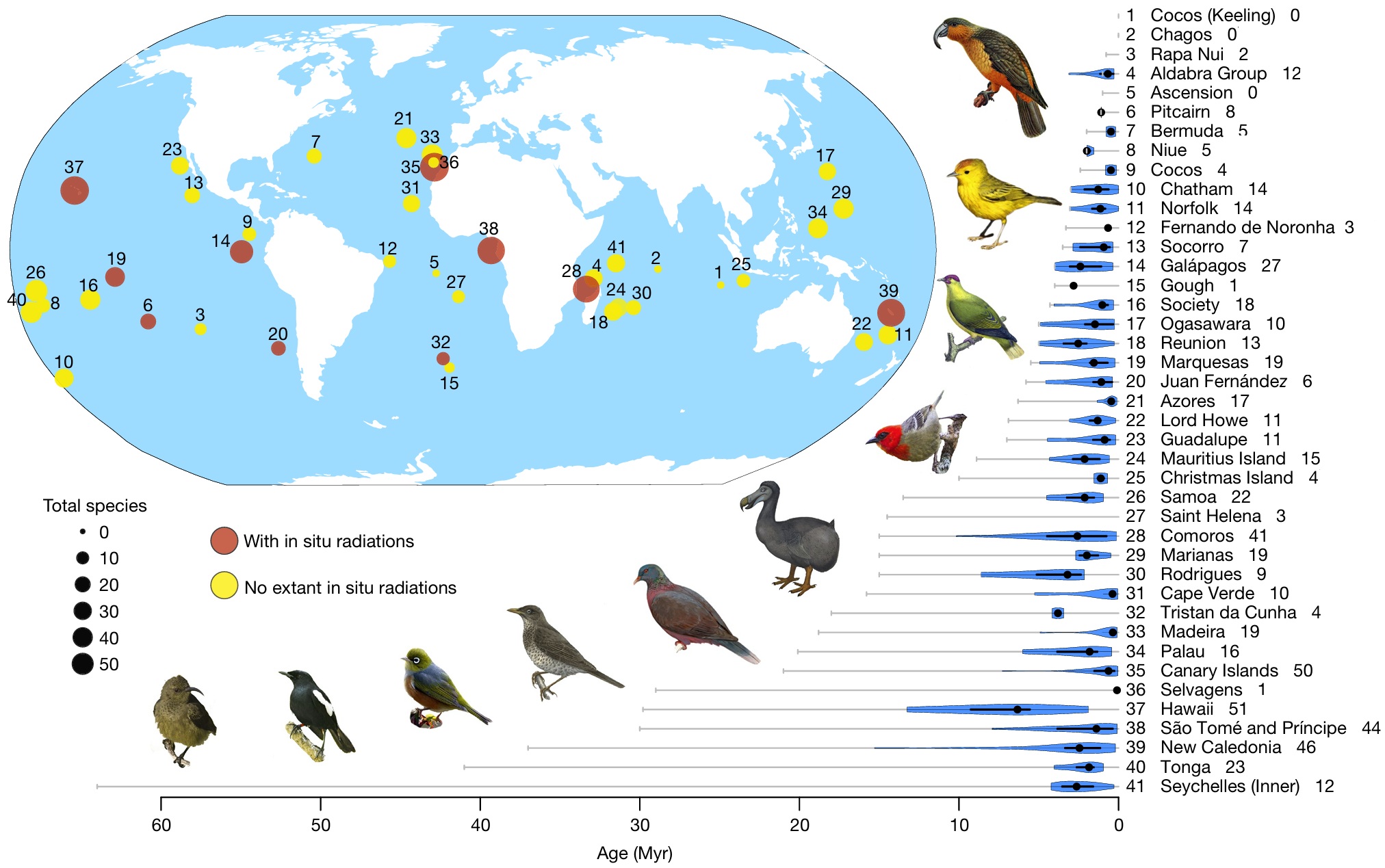
Archipelago and island bird colonization times. Circles show the number of species (both extinct and extant) found in each archipelago (at the time of human arrival).
Numbers on the map correspond to numbers to the left of the archipelago name. Numbers to the right of the archipelago name indicate the number of species on
the archipelago. Colonization times plot: grey horizontal lines indicate archipelago ages. Violin plots (blue) show the distribution of times of colonization of bird species
in each archipelago. Thick black lines inside violin plots indicate the interquartile distance; thin black lines indicate the 95% confidence interval; black dots indicate the median.
Archipelagos with no violin plot or dots are cases where no species were present at the time of human arrival. Birds from left to right: Seychelles Sunbird, Seychelles Magpie Robin,
Silvereye, Príncipe Thrush, Laurel Pigeon, Dodo (extinct), Mauritius Fody, Red-moustached Fruit Dove (extinct), Galápagos Warbler, and Norfolk Kaka (extinct).
Valente et al. (2020) compiled a dataset of birds on islands, based on the terrestrial avifaunas of 41 oceanic archipelagos worldwide (including 596 avian taxa), and applied a new analysis method to estimate the sensitivity of island-specific rates of colonization, speciation and extinction to island features (area and isolation). Their model predictswith high explanatory powerseveral global relationships, including a decline in colonization with isolation, a decline in extinction with area, and an increase in speciation with area and isolation, i.e., rates of speciation were found to increase through an interactive effect of both island size and distance from the nearest mainland, providing a mechanism that limits radiations to archipelagos that are both large and remote. Oceanic Islands
typically support fewer species of land & freshwater birds than equal areas of comparative mainland
obvious influences on bird diversity on islands are age, topography, size, & distance from nearest source of immigrants
importance of size & distance was first quantified by MacArthur & Wilson (1967)
For example, New Zealand (Baker 1991):
Australia & New Zealand are 1200 miles apart
only 65 native species of land & freshwater birds (with an additional 47 species known from fossil remains and 34 species introduced by humans during the past 130 years)
Takahe
avifauna composed largely of elements that dispersed to New Zealand long after the breakup of Gondwanaland
Australian element, not surprisingly, is dominant - prevailing westerly winds regularly deposit Australian species in New Zealand (e.g., 8 species have successfully colonized New Zealand in the past 150 years)
other biogeographical elements include Holarctic (just 3 species) & Archaic (endemic taxa with such a long evolutionary history in New Zealand that no close relatives can be identified elsewhere, e.g., kiwis and moas)
routes of colonization:
Tasman Sea has been the principle route
direct route from the Holarctic (e.g., shorebirds & waterfowl)
Other islands also have (or had) distinct avifaunas, e.g., see Hawaii's Honeycreepers'
New Zealand's Kiwis
The avifauna of the islands of Wallacea includes elements of both Indomalayan and Australian origin (Michaux 2010). Of 513 species of breeding land birds in Wallacea, 300 (58%) are endemics and 213 (42%) are non-endemics that also breed outside of Wallacea. As also found in other island biogeography studies, island geography, especially island area, but also isolation and elevation, largely explain the variation in island species richness and endemism in Wallacea (Dalsgaard et al. 2014).
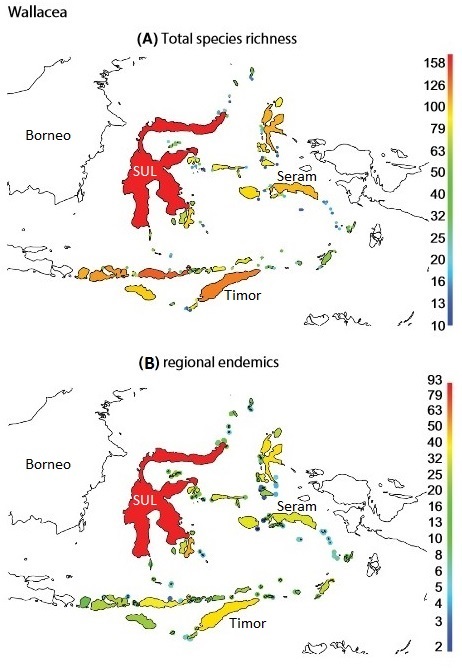
(A) Total species richness on islands in Wallacea. (B) Species richness of regional endemics (only found on islands in Wallacea) in Wallacea.
SUL, Sulawesi (Figure modified a bit from Dalsgaard et al. 2014).
The avifauna of the islands in the tropical Indian Ocean (excluding Madagascar) consists of about 139 species of resident birds (excluding pelagic and introduced species) representing 33 different families of birds. Of these species, 10 species are endemic to the Indian Ocean and 69 are endemic to a single archipelago or island. Most endemic species are in the families Columbidae (pigeons), Psittacidae (parrots), Muscicapidae (thrushes and Old World flycatchers), and Zostertopidae (white-eyes) (Adler 1994). Unfortunately, several of the 139 species of resident birds are now extinct.
The presence of relatively few endemic species of birds on the islands of the tropical Indian Ocean is due to the almost complete absence of any intra-archipelago speciation, likely because the Indian Ocean archipelagos have too few of the large islands needed for such speciation. In addition, some families of birds that are widespread in the Afrotropical and Indomalayan realms, such as Alcedinidae (kingfishers) and Sturnidae (starlings), are either absent or poorly represented on islands in the tropical Indian Ocean. For some families, e.g., kingfishers, this absence may be due the absence of suitable habitat (rivers, streams, or lakes). For other families, reasons for their absence is less clear (Adler 1994).
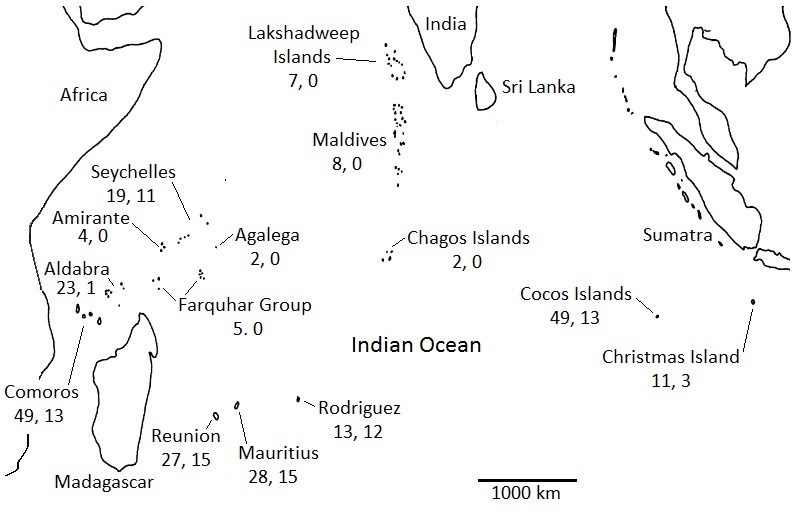
Avifaunas of islands in the Indian Ocean. The first number below each island’s and group of island’s name is the
number of species of resident breeding birds; the second number is the number of endemic species (Figure modified from Adler 1994).
The islands of the Pacific originated in a variety of ways, including as linear chains of volcanic islands, fragments of continental crust, and obducted portions of adjoining tectonic plates (Neall and Trewick 2008).
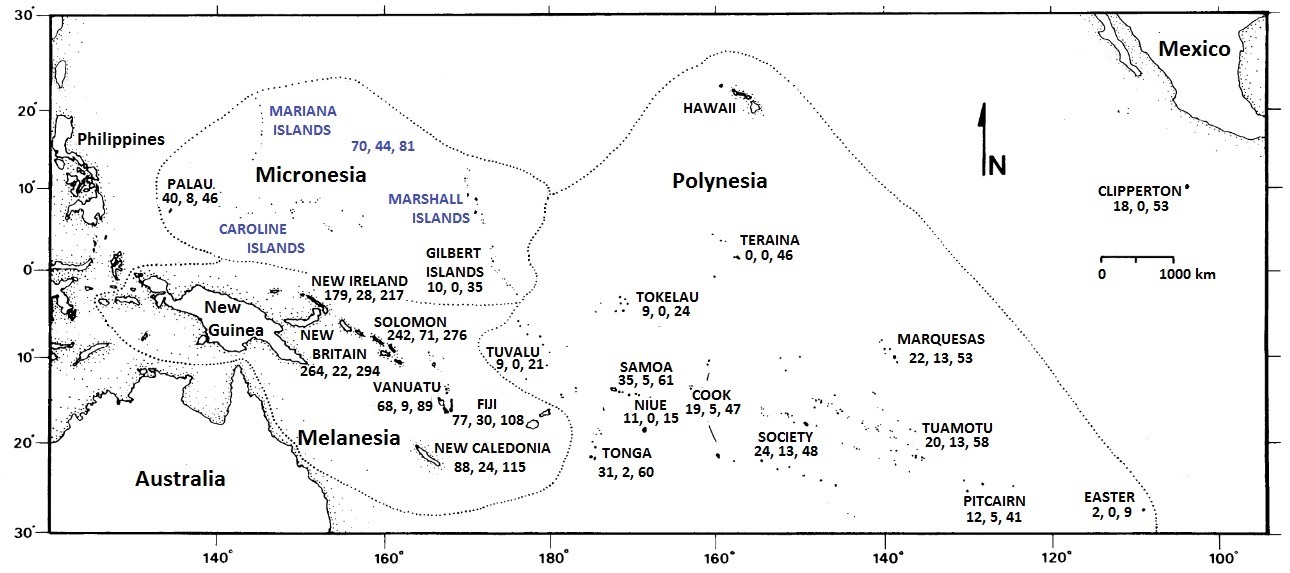
Avifaunas of islands in the Pacific Ocean. Numbers below the names of islands indicate, in order, the number of
breeding land birds, number of endemic species, and total number of species including seabirds (Figure modified from Steadman 1995).
The Hawaiian archipelago consists of 45 islands extending across 2450 km of the central Pacific Ocean and is the most remote archipelago on the planet (Juvik and Austring 1979). The Hawaiian Islands once had at least 111 native species of birds, almost all endemic, before humans (Polynesians) arrived about 1000 years ago. Of those 111 species, 55 became extinct after arrival of the Polynesians and are now known only as fossils, and 19 more extinctions occurred after arrival of Europeans (Scott et al. 2001). Of the remaining 37 extant endemic species, 33 are at some risk of extinction. In addition to these endemic species, at least 22 species of seabirds breed on the Hawaiian Islands, and another 54 species of landbirds have been introduced by humans and now breed in Hawaii.
All of Hawaii’s endemic species of birds are thought to have originated from just 20 colonization events, and the many species of honeycreepers (originally more than 50, just 17 remain) are thought to be derived from a single ancestor (Berger 1981, Olson and James 1982). Based on analyses of mitochondrial genomes, Lerner et al. (2011) suggested that Hawaiian honeycreepers are a sister taxon to Eurasian rosefinches (Carpodacus) and that the ancestor of the honeycreepers arrived in Hawaii about 6-7 million years ago. In addition to the honeycreepers, the endemic Hawaiian avifauna includes (or included) several waterfowl, two raptors, a crow (extinct in the wild, but some still live in captivity), and representatives of Old World flycatchers, honeyeaters, and thrushes (Jacobi and Atkinson 1995). The ancestors of these and other endemic species arrived in the Hawaiian Islands beginning about 5 million years ago.
Estimated arrival times (millions of years ago, Mya) of the ancestor of several species and taxa of birds in the Hawaiian Islands
(Price and Clague 2002; except Hawaiian honeycreepers from Lerner et al. 2011).
Lineage |
Age (Mya) |
No. of species |
Hawaiian honeycreepers (Fringillidae: Drepanidinae) |
7.2 |
~50 |
Laysan Duck (Anas laysanensis) |
<5 |
1 |
Hawaiian Crow (Corvus hawaiiensis) |
<4.2 |
1 |
Flightless Anseriformes (‘moa-nalos’) |
<3.6 |
4 |
Hawaiian thrushes (Myadestes spp.) |
<3.35 |
5 |
Flightless rail (Porzana sandwichensis) |
<2.95 |
1 |
Flightless ibises (Apteribis spp.) |
<1.6 |
2 |
Hawaiian duck (Anas wyvilliana) |
<1.5 |
1 |
Flightless rail (Porzana palmeri) |
<1.05 |
1 |
Hawaiian geese (Branta spp.) |
<1 |
3 |
Hawaiian Black-necked Stilt (Himantous mexicanus knudsenii) |
<0.75 |
1 |
Hawaiian Hawk (Buteo solitarius) |
<0.7 |
1 |

Some of Hawaii’s endemic species of birds. (A) a male ‘Akeke’e (Loxops caeruleirostris), (B) a female O’ahu ‘Elepaio (Chasiempis sandwichensis)
feeding nestlings, (C) ʻĀkohekohe (Crested Honeycreeper, Palmeria dolei), (D) a male ‘I’iwi (Vestiaria coccinea), (E) Puaiohi (Myadestes palmeri)
at its nest, (F) a male Palila (Loxioides bailleui), (G) a male Akiapōlā’au (Hemignathus wilsoni), and (H) a Nēnē (Branta sandvicensis)
with young (Figure made of photos from VanderWerf 2012).
The islands in the Atlantic Ocean off the coasts of southern Europe and northern Africa are collectively referred to as Macaronesia. This region has 31 principal volcanic islands that are grouped into five archipelagos: Azores, Madeira, Selvagens, Canary Islands, and Cape Verde (Figure below). These archipelagos vary in their distance from the mainland (range = <100 km for the Canary Islands and 1365 km for the Azores), latitude, and age (range = 0.25 – 29 million years) (Hazevoet 1995, Geldmacher et al. 2001, Azevedo and Ferreira 2006). The taxonomic affinities of native bird species suggest that the origin of the extant terrestrial avifauna is largely from the western Palaearctic mainland, but the Cape Verde archipelago, because of its lower latitude and proximity to Africa, has species related to mainland Africa (Hazevoet 1995, Martín and Lorenzo 2001, Clarke et al. 2006). In addition, the avifaunas of the four largest archipelagos (excluding Selvagens) have all reached an equilibrium, with species richness reaching a plateau because rates of colonization and speciation are similar to extinction rates (Valente et al. 2017).
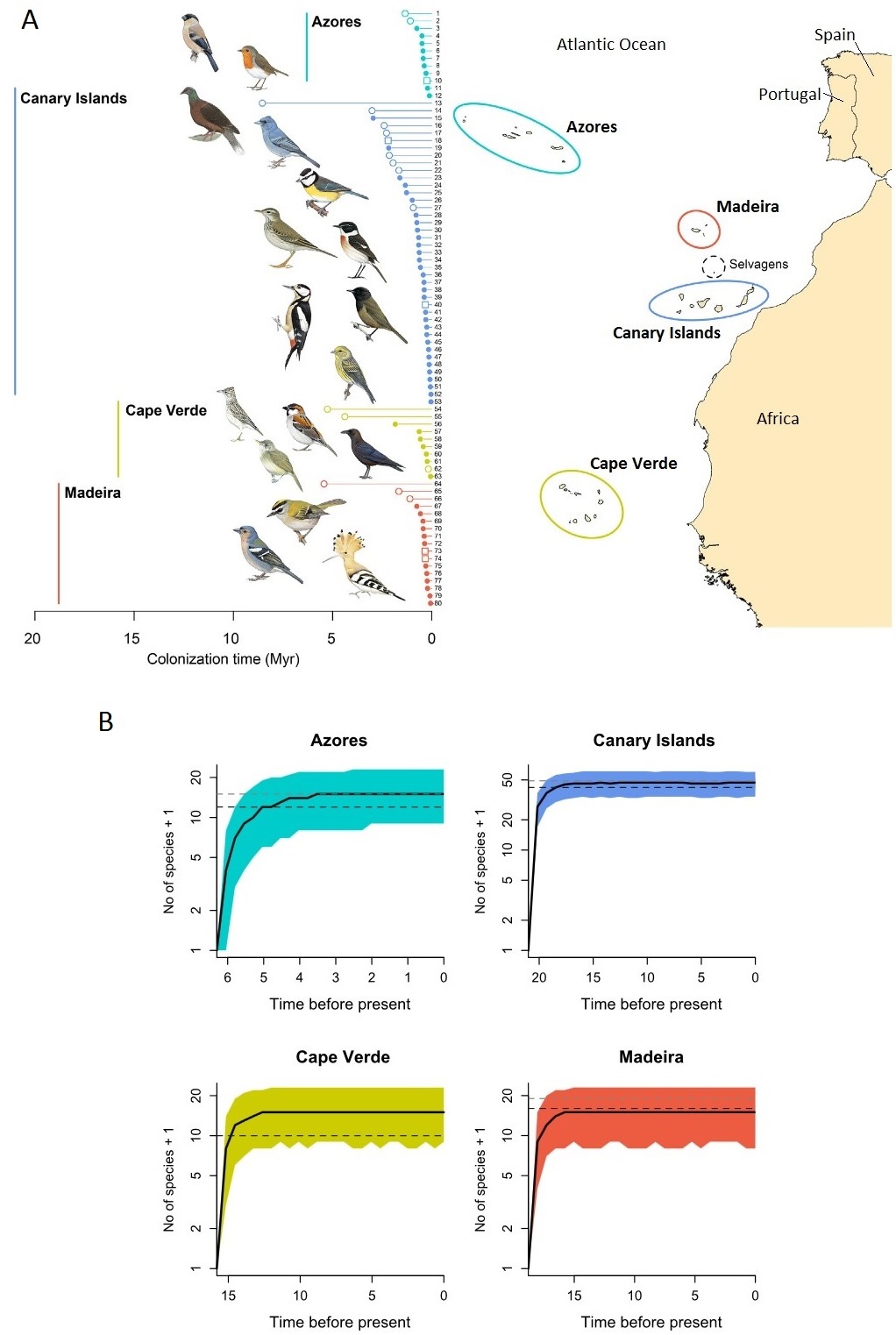
(A) Colonization times of Macaronesian species of landbirds and a map of Macaronesia. The vertical lines to the left indicate the maximum geological ages of the archipelagos.
Filled circles, non-endemic species; unfilled circles, endemic species; unfilled squares, Macaronesian endemic. The Selvagens Islands, although important breeding areas for several species of seabirds,
have only two resident landbirds: Berthelot’s Pipit (Anthus bertheloti) and Common Kestrel (Falco tinnunculus). (B) Estimated n umbers of species over time in each of the archipelagos.
Black lines are estimated median values and the colored areas show the 2.5th–97.5th percentiles. Gray dashed line, pre-human diversity; black dashed line, present diversity (excluding extinct species).
Time before present is in millions of years (Figures from Valente et al. 2017).
The West Indies consist of three groups of islands: (1) the Greater Antilles, including Cuba, Hispaniola, and Puerto Rico, are old fragments of continental crust that originally formed as islands closer to present-day Central America and carried to their present locations by plate movements, (2) the Bahama islands are low-elevation islands located north of the Greater Antilles and southeast of Florida, and (3) the Lesser Antilles were created by a series of volcanoes, some still active, where the South American Plate is being subducted under the Caribbean plate. Bird species richness of the West Indies is low compared to the nearby continents, likely because of the limited dispersal abilities of many avian taxa (Ricklefs and Bermingham 2001). Water barriers reduce the likelihood of colonization by birds that rarely move beyond forest borders like most tropical suboscine passerines and woodpeckers (Terborgh 1973, Terborgh et al. 1978).
Birds of Cuba
Of the 243 breeding species in West Indies, 176 (72%) are endemics and 67 (28%) non-endemics (Dalsgaard et al. 2014). The main pathways of immigration into the islands have been from the south (the northern coast of South America and through the Lesser Antilles), from the west (Central America and into the Greater Antilles), and from the north (Florida). Although South America contains a larger species pool of potential immigrants and the Lesser Antilles are close to the coast of South America, more species of birds in the West Indies originated in North and Central America (Bond 1963). However, species that have invaded the West Indies from South America have tended to move further north than species that emigrated from North and Central America. As is typically the case, the larger islands in the West Indies have more landbird species and more endemic species of birds.
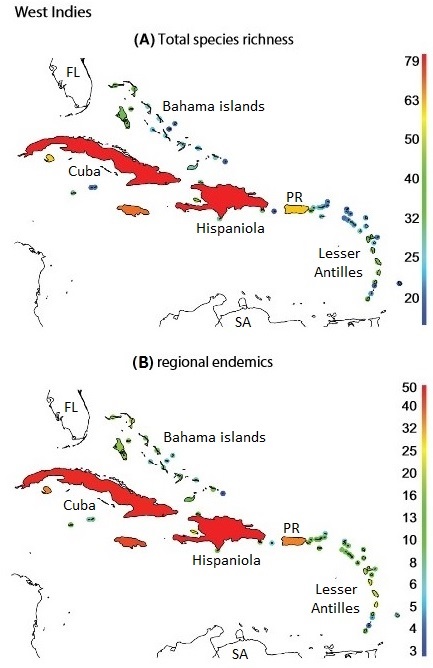
A) Total species richness on islands in the West Indies. (B) Species richness of regional endemics, i.e., only found on islands
in the West Indies. FL, Florida; PR, Puerto Rico; SA, South America (Figure modified from Dalsgaard et al. 2014).
Seminar - Challenges and opportunities for Caribbean endemic bird conservation
The islands of the Southern Ocean can be divided into older islands with a continental origin (e.g ., Falkland Islands, South Georgia, Auckland and Campbell Islands), and younger, volcanic islands (e.g., most islands in the southern Indian Ocean). Islands in the Southern Ocean have cool, wet, and windy climates. The islands are covered in tundra vegetation, ranging from scattered dwarf plants to grasslands, herbfields, and bogs. Vegetation in coastal areas includes tussock grasses, graminoids, and herbs; more upland areas are often covered with cushion-forming angiosperms and mosses. Vascular plant species richness ranges from zero (Bounty Islands) to 188 (Auckland Islands), and insect species richness varies from six (McDonald Island) to more than 200 (Auckland Islands) (Bergstrom and Chown 1999).
With the exception of the Falkland and Chatham islands, most islands in the Southern Ocean have few or no breeding landbirds. In addition, the Southern Ocean islands have few endemic species. The few breeding songs generally show affinities to the nearest mainland areas. In contrast to the few breeding landbirds, the Southern Ocean islands are the primary nesting areas for numerous species of seabirds. The productivity of the southern oceans provides abundant food for breeding seabirds.
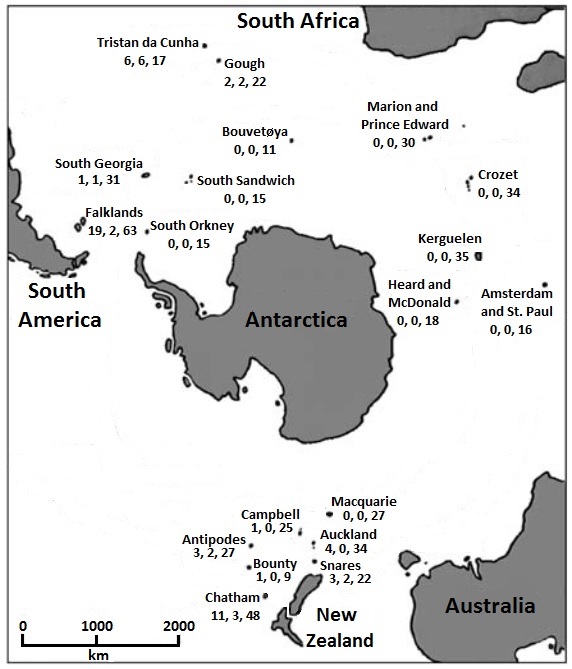
Islands of the Southern Ocean. Numbers below the names of the islands indicate the number of breeding landbirds, number of endemic species,
and total number of breeding birds. Most birds breeding on these island are penguins, albatrosses, petrels and prions,
shearwaters, storm-petrels, skuas, and gulls and terns (Numbers from Shirihai 2007; Figure modified from Angel et al. 2009).
Pacific birds rewrite evolutionary textbook-- Evolutionary biologists have long assumed that species in island-rich areas
such as the Pacific Rim
diverge and evolve in a series of 'stepping-stone'
dispersals from one island to the next. But a new analysis of the family
tree of tropical Pacific
birds suggests that this cherished assumption is
not true.
Filardi and Moyle (2005) analysed DNA from a host of bird
species from the family
Monarchidae, commonly called Monarch
flycatchers. They found that the species seem to have colonized the
area not in a series of separate
stepping-stone excursions from continental
land masses, but in a single, even spread across all Pacific archipelagos.
The researchers support their
conclusion by pointing out that, of the six
different species groups that are confined to a single island chain, all are
embedded within a group, or
genus, called Monarcha. This genus is spread
across the entire tropical Pacific, showing that all of these remote species
arose from a single lineage.
What's more, the authors point out, some
species show evidence of having recolonized continental areas such as
Australia - contrary to the previous
idea that that species colonize island
chains only by moving away from continents.
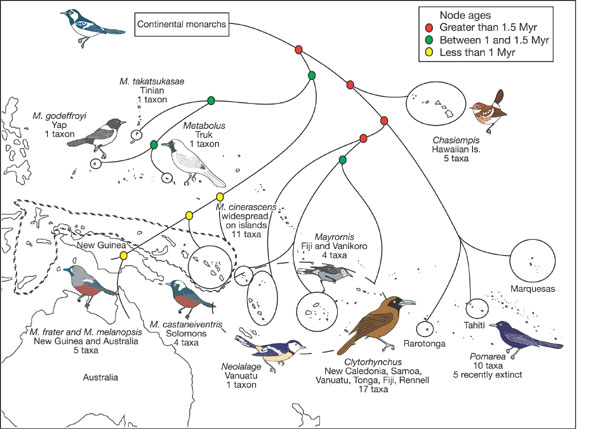
Bird drawings are roughly to scale and illustrate some of the marked morphological differentiation between island endemics that
has complicated the evolutionary interpretation of the group. Colored circles indicate three classes of node age. Branch lengths are
not to scale. Large hatching outlines the distribution of Clytorhynchus and small hatching outlines the range of Monarcha cinerascens.
|
||
| Ancient
DNA Tells Story of Giant Eagle Evolution --
For reasons not entirely clear, when animals make their way to isolated
islands, they tend to evolve relatively quickly toward an outsized or
pint-sized
version of their mainland counterpart. One avian giant was found in New
Zealand: Haast's eagle (Harpagornis moorei), with a
wingspan
up to 3 meters. Though Haast's Eagle could fly, its body mass
(10–14
kilograms) pushed the limits for self-propelled flight. As extreme
evolutionary
examples, large island birds can offer insights into the forces and
events
shaping evolutionary change. Bunce et al. (2005) compared ancient
mitochondrial
DNA extracted from Haast's Eagle bones and found that the extinct
raptor
underwent a rapid evolutionary transformation that belies its kinship
to
some of the world's smallest eagle species. Their analysis places
Haast's
Eagle in the same evolutionary lineage as a group of small eagles in
the
genus Hieraaetus. Surprisingly, the genetic distance separating
the giant eagle and its smaller cousins from their last common ancestor
is relatively small.
Using molecular dating techniques, Bunce et al. (2005) proposed a divergence date of 0.7–1.8 million years ago. The increase in body size—by at least an order of magnitude in less than 2 million years—is remarkable, Bunce et al. (2005) argue, because it occurred in a species still capable of flight. The absence of mammalian competitors may have precipitated the rapid morphological shift. Haast's eagle, the authors wrote, “represents an extreme example of how freedom from competition on island ecosystems can rapidly influence morphological adaptation and speciation.” -- Source: (2005) Ancient DNA Tells Story of Giant Eagle Evolution. PLoS Biol 3(1): e20. |
Avifaunas of the various zoogeographic realms represent a
mixture
of species of various ages & origins. The mobility of many (but not
all) birds, plus their proclivity to migrate, has allowed the movement
of some groups between realms - some very long ago, others more
recently.
And, once a species arrives in a new area, adaptive radiation often
generates
new species. This combination of avian mobility (or lack thereof) and
adaptive
radiation has created avifaunas that represent complex mosaics
(Gill  1995).
Briefly, the avifaunas of the major zoogeographic realms can be
described
as follows:
1995).
Briefly, the avifaunas of the major zoogeographic realms can be
described
as follows:
Palearctic
many genera shared with the Nearctic, Ethiopian, & Oriental regions indicating much dispersal
geographic 'barriers' between these regions include:
the Sahara desert (Ethiopian)
the Himalayas (Oriental) (viewed from space in photo to the right)
the Bering Sea (Nearctic)
relatively low species richness, probably due to cold climate & large areas of homogeneous habitat (taiga)
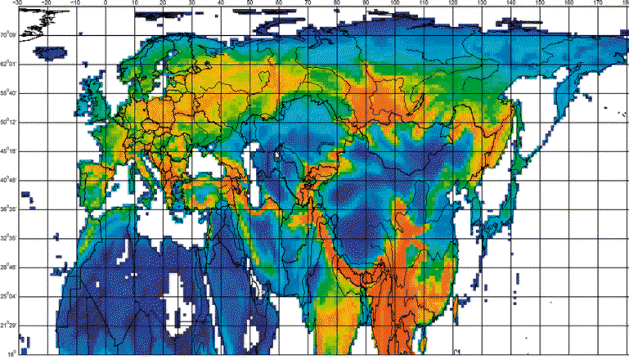
Map of species richness among equal area grid cells in the full data set (3,036 taxa) of Palearctic songbirds.
Maximum richness shown in deep rufous and minimum in dark blue. Scores grouped into 32 color-scale classes of
approximately equal frequency. Classes range from 1–3 species per grid cell (blue) to 128–307 (rufous); also a single grid cell
with maximum score (316 species, in northeast Myanmar [Burma]) in red (Roselaar et al. 2007). Most 'hotspot' regions are located
in mountainous regions.
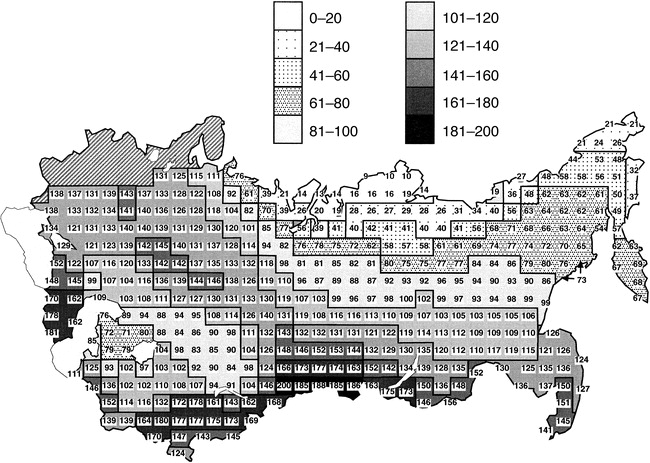
Geographic variation in the richness of bird species in the former USSR. The hatched area in the upper left represents an area classified as the western Palearctic (Hawkins et al. 2003).

Examples of songbirds found in the Palearctic realm. (A) Long-billed Pipit, Anthus similis (Photo by Ian Boustead), (B) Indian Blue Robin, Luscinia brunnea
(Photo by Niranjam Sant), (C) White-throated Redstart, Phoenicurus schisticeps, (D) White-capped Water Redstart, Chaimarrornis leucocephalus,
(E) Black-browed Tit, Aegithalos bonvaloti, (F) Chestnut Thrush, Turdus rubrocanus, (G) Temminck’s Lark, Eremophila bilopha (Photo by Augusto Faustino),
(H) Plain-backed Thrush, Zoothera mollissima (Photos C, D, E, and F by John and Jemi Holmes) (Figure made from photos from Roselaar 2006).
The Western Palearctic Avifauna (Blondel and Mourer-Chauviré 1998) -- Compared with that of the two other large forested regions of the Northern Hemisphere (eastern North America and eastern Asia), the bird fauna (and particularly forest avifauna) of the western Palearctic is relatively species poor. Controlling as much as possible for the size of areas, Mönkkönen and Viro (1997) have shown that the western Palaearctic (including North Africa) has 50% fewer forest-associated bird species than eastern Asia and 40% fewer than eastern North America. Europe is similar in size to China (9 500 000 km2, including Manchuria), but European species  richness (about 500 bird spp.) is only about half that reported for China. In the western Palaearctic, less than half of the terrestrial avifauna is associated with forest compared with two-thirds in eastern North America and eastern Asia.
richness (about 500 bird spp.) is only about half that reported for China. In the western Palaearctic, less than half of the terrestrial avifauna is associated with forest compared with two-thirds in eastern North America and eastern Asia.
Large and repeated habitat changes during the Pleistocene have undoubtedly accelerated the extinction rates of birds of the western Palearctic, just as they did for plants and large mammals in North America. Comparative analyses using molecular phylogenies of 11 lineages of North American songbirds also indicate a general decrease in rates of species diversification during the Pleistocene. However, molecular evolutionary trees cannot distinguish between reduced speciation rates (e.g. as a consequence of progressive niche filling through evolutionary time) and higher extinction rates as the cause of net changes in diversification rates.
Assuming similar rates of speciation over the whole Northern Hemisphere during the Pleistocene, the relative impoverishment of the western Palearctic bird fauna (compared with the two other regions of the Northern Hemisphere) could be caused by higher extinction rates in the western Palearctic, because there was no connection with the tropical biomes further south. In contrast to those of the western Palearctic, the extant bird faunas of both northeastern North America and eastern Asia share many species in common with southern tropical regions. For example, 76% of passerine species breeding in North America have either conspecific populations or congeneric species that breed in the Neotropics. In eastern Asia, two orders and nine families are tropical, whereas Afro-tropical influence in the fauna of the western Palaearctic is negligible.
The most likely explanation for these differences is that the biota of both eastern North America and eastern Asia remained connected to the tropics over the whole Tertiary–Quaternary, making it possible for a continuous interchange between tropical and temperate avifaunas. In contrast, the massive east–west oriented barriers (mountain ranges, seas and large desert belts) ‘trapped' biota of the western Palaearctic in their southern refugia during glacial periods, preventing them from finding refuge in tropical regions further south, and preventing tropical species from colonizing northern regions. Since the Pliocene, the Sahara desert has never been sufficiently forested to provide a dispersal corridor for forest birds between the Afro-tropics and Eurasia.
Nearctic
Species richness, compared to most other realms, is relatively low. Contributing factors include:
Geographic variation in the richness of breeding terrestrial bird species in the Nearctic and northern Neotropics. The heavy dashed lines in southern Mexico distinguish cells classified as Nearctic and cells classified as Neotropical. Numbers represent the number of bird species in each cell (Hawkins et al. 2003).
| Deciduous forest may have barely existed in the southern USA at the Last Glacial Maximum (LGM). Contrary to most of the earlier interpretations, it now seems inappropriate to show any significant area of deciduous forest in the eastern US for the LGM. Current work by S. Jackson (Northern Arizona University), Eric Grimm (Illinois State Museum), and Bill Watts suggests that even in the south there was very little broad-leaved forest at the LGM, with perhaps only isolated pockets of deciduous forest that were surrounded by coniferous vegetation. These deciduous forest pockets at the LGM may have been confined to moister microsites, such as stream gullies (Ritchie 1982). For example, spruce forest has been found to have been growing in Louisiana at the LGM. Tallis (1990) also maps a south-eastern pine forest that extending from about 33°N down to the Gulf coast and northern Florida, and suggests that it would generally have been fairly open in structure. Source: http://www.soton.ac.uk/~tjms/namerica.html |
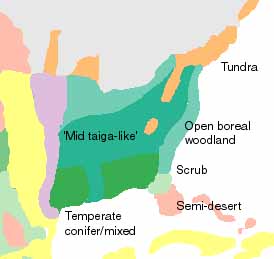 Source: http://www.esd.ornl.gov/projects/qen/NAL2215.gif & http://www.esd.ornl.gov/projects/qen/nercNORTHAMERICA.html |
avifauna appears to be composed of six elements (Mayr 1946):
'Cosmopolitan' - 'mobile' groups with world-wide distributions, including hawks (Accipitridae), swifts (Apodidae), woodpeckers (Picidae), & nightjars (Caprimulgidae)
'Panboreal' (or Holarctic) - groups found throughout the Palearctic-Nearctic, including loons (Gaviidae) & some shorebirds
'Pan-American' (or Neotropical) - groups that originated in South America, including ,tyrant flycatchers (Tyrannidae), and tanagers (Thraupinae)
'Pantropical' - groups with undetermined origins in the southern continents, including parrots (Psittacidae)
'Old World' (or Palearctic):
early colonizers, including cranes, pigeons, cuckoos, owls, crows, & some thrushes
later colonizers, including kingfishers, jays, titmice, & some cardueline finches
recent colonizers, including the barn owl, some swallows, some thrushes, kinglets, & shrikes
'Nearctic' - groups that likely originated in North America, including grouse, turkey, wrens, waxwings, & warblers (Parulidae)
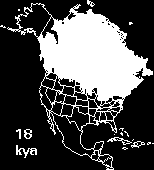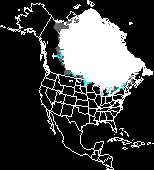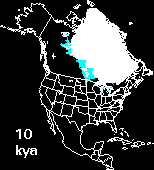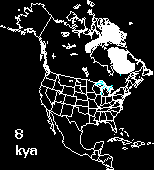
flickers (Colaptes spp.; Mayr 1942)
juncos (Junco spp.; see 'Postglacial range expansion drives the rapid diversification of the genus Junco' & bluebirds (Sialia spp.) (Rand 1948)
warblers (Setophaga & Vermivora spp.; Mengel 1964)
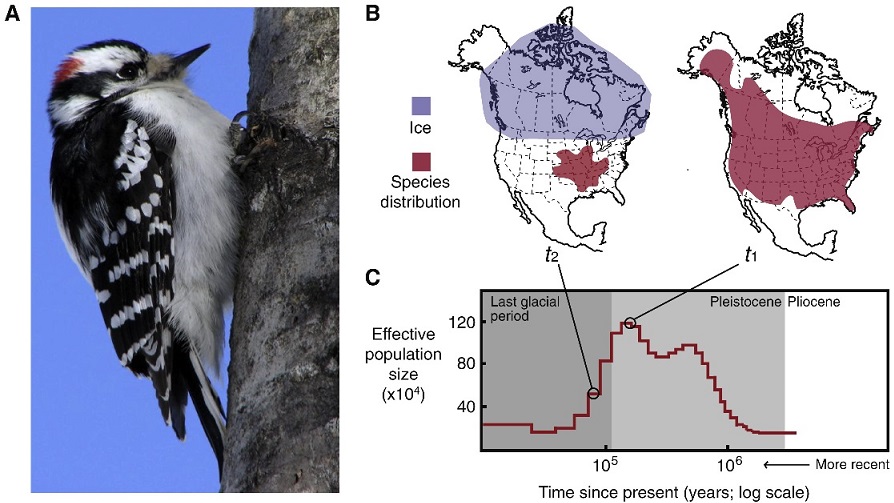
(A) Male Downy Woodpecker (Picoides pubescens) (photograph by Peter de Wit; Wikimedia Commons).
(B) A possible scenario for dynamic range shifts at high latitudes responding to glaciations by contracting (t2) and
then expanding (t1) during more favorable times. (C) Variation in historical population size for Downy Woodpeckers
through time (From Toews 2015).
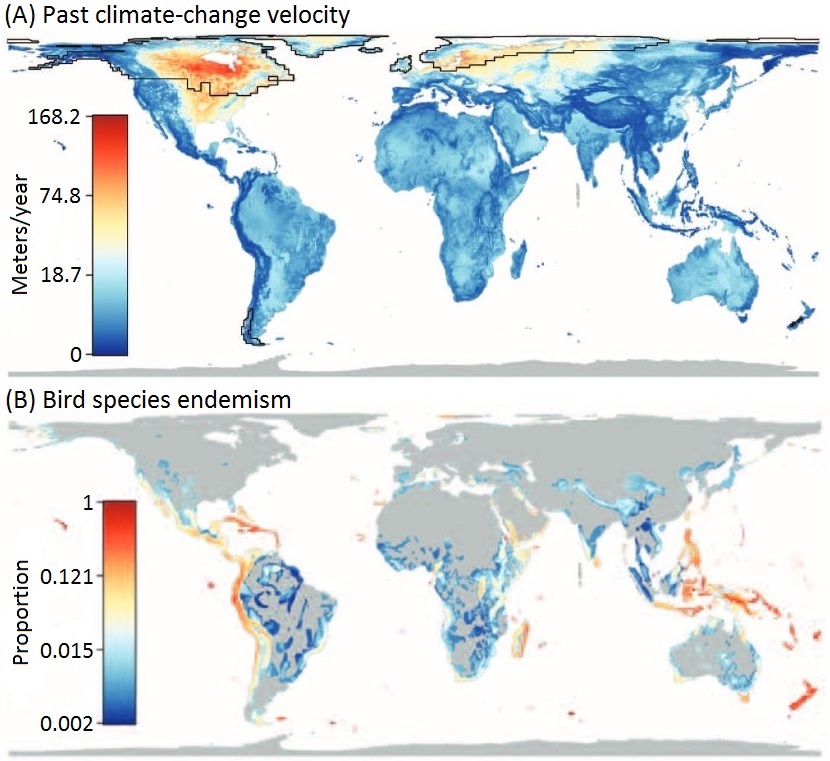
Few endemic species of birds are found in the Nearctic and the Palearctic. One important reason for this is that climates in
the northern hemisphere have changed dramatically since the Last Glacial Maximum (LGM; 21,000 years ago). (A) Climate-change velocity
(rate of movement of climatic conditions over the Earth’s surface) since the LGM. Velocity is highest in northeastern
North America (Nearctic) and north-central Eurasia (Palearctic), in and near the areas that were glaciated (delimited by black lines).
(B) Proportion of bird species that are endemic throughout the world. Areas where climate-change velocities have been highest are also the
areas with the fewest endemic species, likely due to species extinctions as well as incomplete range filling. Areas with greater
proportions of endemic species are also the areas where climates have exhibited little or no changes since the LGM. Note also
that many of the areas with high proportions of endemic species are islands
(Figures from Sandel et al. 2011).
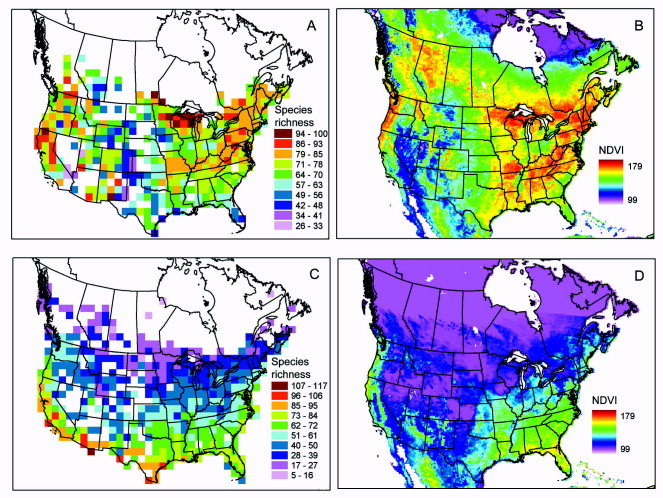 A, Summer pattern of species richness across North America on the basis of expected number of species over three randomly chosen Breeding Bird Survey routes per 20,000-km2 grid cell. B, Spatial pattern of normalized difference vegetation index (NDVI) in June at 1-km resolution. C, Winter pattern of species richness across North America on the basis of expected number of species over one randomly chosen Christmas Bird Count circle per 20,000-km2 grid cell. D, Spatial pattern of NDVI in December at 1-km resolution (From: Hurlbert and Haskell 2003). |
Ethiopian (Afrotropical)
strong affinities with the Oriental realm; many Palearctic species winter south of the Sahara desert
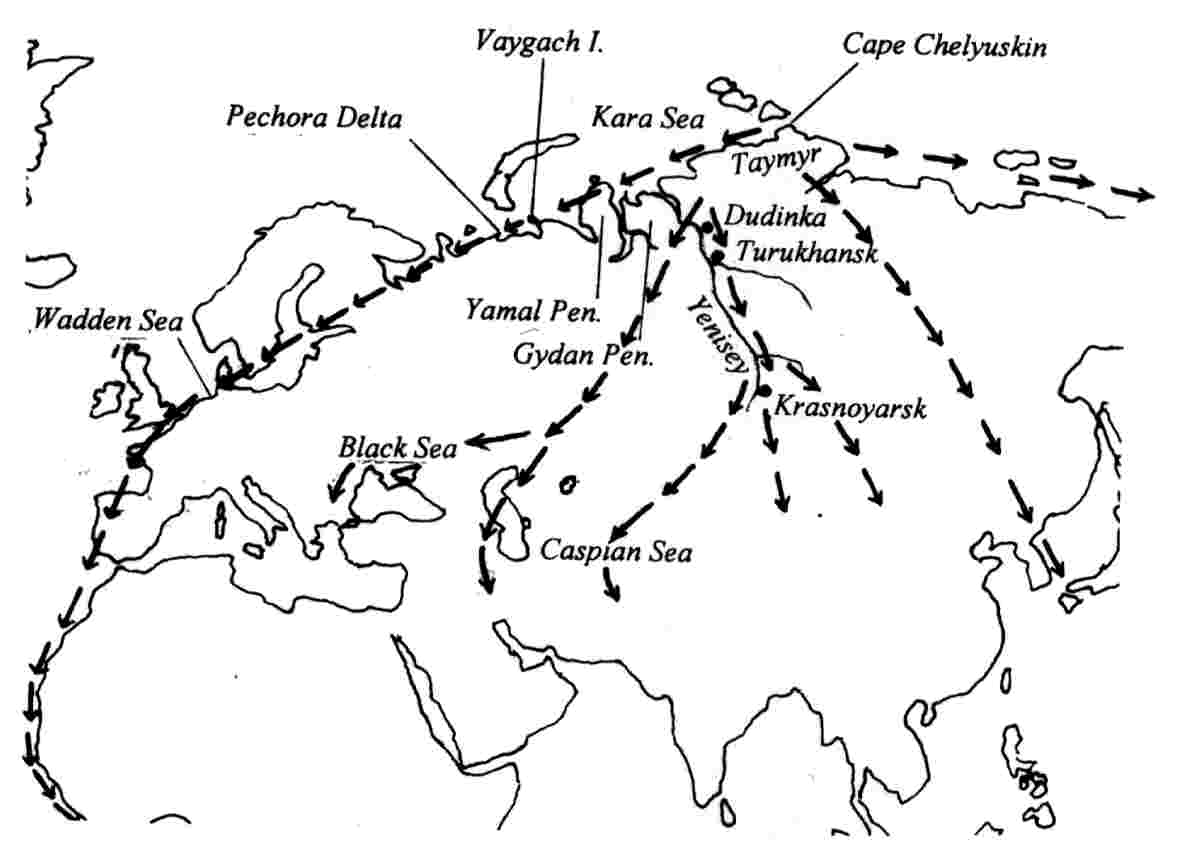
similar latitude to Neotropical realm, but fewer than half as many species:
less tropical forest & generally much drier than the Neotropics
few mountain ranges
endemic families generally contain relatively few species, e.g.:
1 ostrich (Struthionidae)
1 hamerkop (Scopidae)
1 shoe-billed stork (Balaenicipitidae)
1 secretarybird (Sagittariidae)
6 mousebirds (Coliidae)
8 woodhoopoes
44 helmet shrikes (Prionopidae)
23 turacos (Musophagidae)
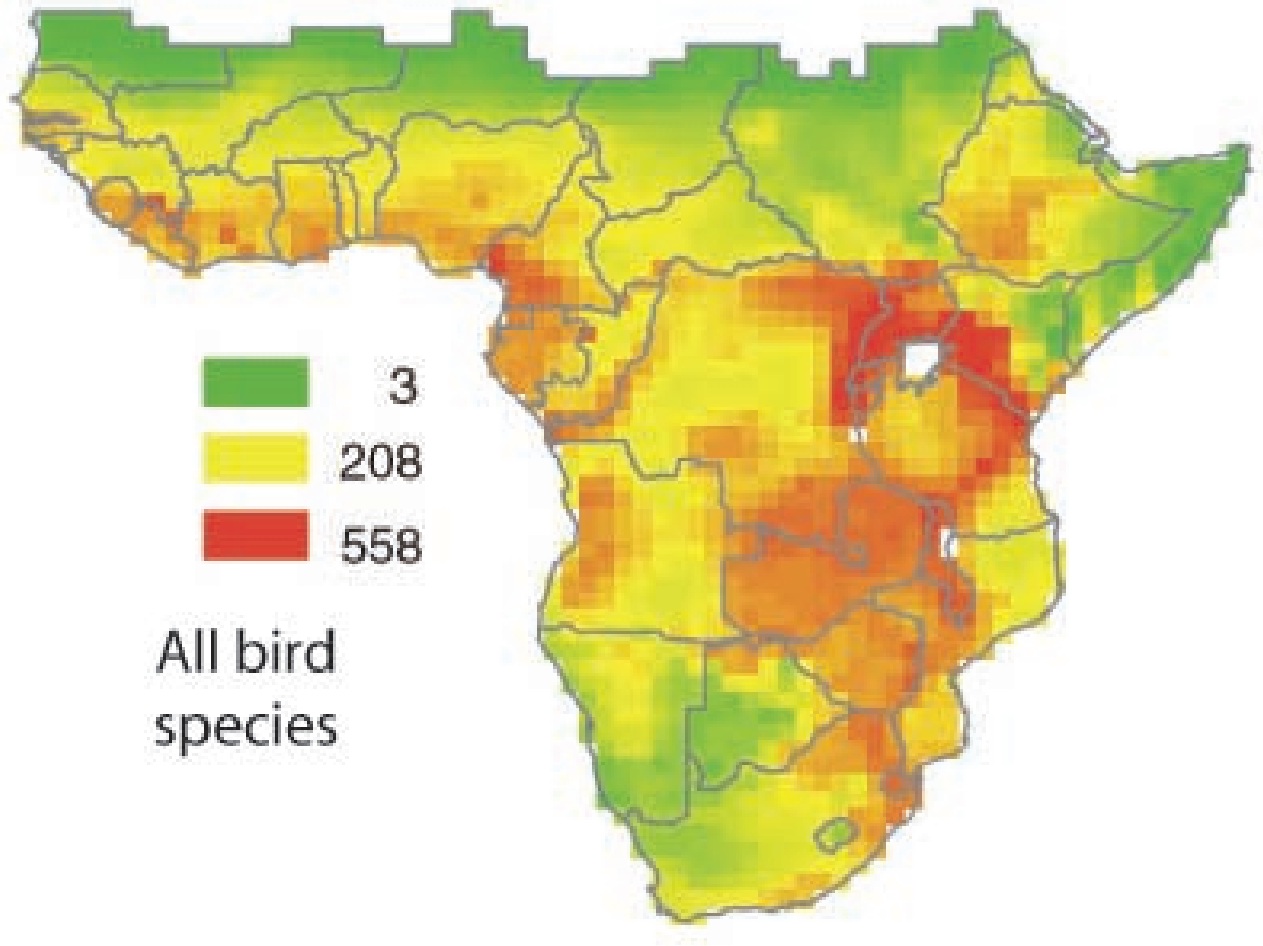
Species richness for 1599 species of birds endemic to Africa (Figure from: Colwell et al. 2009).
Species richness of endemics (species globally restricted to sub-Saharan Africa) peaks along the
mountains and adjacent lowlands of eastern and southern Africa. Species richness is also relatively
high in the isolated mountains in central and western Africa and the lowlands of the northeastern
Congo Basin (de Klerk et al. 2002).
Birds of Africa
Ancient African birds
Lake Nakuru's flamingoes
Turacos are uniquely African, with 23 species recognized by most authorities. The forest-living species are undoubtedly among the most spectacularly colorful of birds, while the savanna-dwellers (known as "go-away birds" because of a well-known call, a harsh "kay-waaay") are predominantly grey in plumage. In South Africa, these birds are better known as louries. All are frugivores, specializing in fruit (particularly figs) and sometimes feeding on leaves, buds and flowers. They are usually gregarious, and frequently associate with parrots, hornbills and barbets, with individuals and groups often returning day after day to the same fruiting tree, until the crop is exhausted. |
 White-bellied Go Away Bird |
Oriental (Indomalayan)
closest affinities to tropical Africa, but linkages to Palearctic & Australian faunas as well. The Himalayas represent a boundary between Oriental & Palearctic regions; water separates the Oriental & Australian regions.
only one endemic family (Irenidae), but another nearly so (Eurylaimidae); pheasants (Phasianidae) have undergone considerable radiation
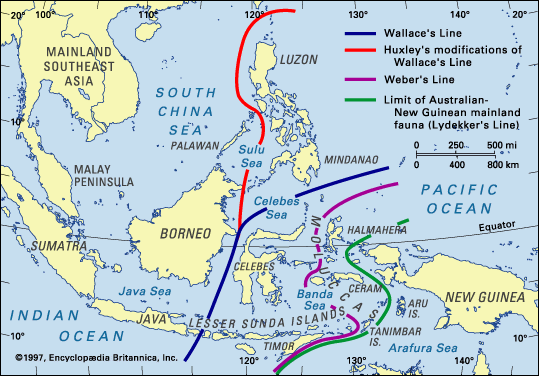
boundary between Oriental & Australasian realms is Wallace's Line; more recent work indicates that the 'line' runs roughly between Celebes & the Moluccas (~600 km east of Wallace's Line & known as Weber's Line
| Birds & the Beetles --
Batrachotoxins are
neurotoxic steroidal alkaloids first isolated from a Colombian
poison-dart
frog. In 1992, the toxic principle from feathers and skin of birds in
the
genus Pitohui, endemic to New Guinea, was isolated and, remarkably,
proved
to be mainly a batrachotoxin (BTX). BTXs were later found in New Guinea
toxic birds of the genus Ifrita. On a weight basis, the batrachotoxins
are among the most toxic natural substances known, being 250-fold more
toxic than strychnine. It is believed that such toxins provide some
protection
against the birds' natural enemies, such as parasites and predators,
including
humans. Neither poison-dart frogs or birds are thought to produce the
toxins
de novo, but instead they likely sequester them from dietary sources.
Dumbacher
et al. (2004) described the presence of high levels of batrachotoxins
in
a little-studied group of beetles, genus Choresine (family Melyridae).
These small beetles and their high toxin concentrations suggest that
they
might provide a toxin source for the New Guinea birds. Stomach content
analyses of Pitohui birds revealed Choresine beetles in the diet, as
well
as numerous other small beetles and arthropods. The family Melyridae is
cosmopolitan, and relatives in Colombian rain forests of South America
could be the source of the batrachotoxins found in the highly toxic
Phyllobates
frogs of that region.
The Blue-capped Ifrita (Ifrita kowaldi; shown to the right) is called Nanisani by local villagers. Nanisani is also the word used to describe the tingling and numbing sensation of the lips and face that results from contacting both beetles and bird feathers (Photo Source: Smithsonian National Zoological Park). |
 A Choresine pulchra beetle with an insert showing glandular vesicles.
|
Jack Dumbacher and Hooded Piohuis
Australasian
Long history of isolation from other land masses
16 families endemic (or nearly so); of 308 passerine species, 80% are 'old endemics' belonging to lineages that likely originated & radiated in Australia & New Guinea
Some evidence that the parvorder Corvi began an evolutionary radiation in Australia about 55-60 million years ago (Sibley and Ahlquist 1985) & gave rise to:
Superfamily Menuroidea
Australian treecreepers (Climacteridae)
lyrebirds (Menuridae)
bowerbirds (Ptilonorhynchidae) (check this video about the Vogelkop Bowerbird)
Superfamily Meliphagoidea
fairywrens & grasswrens (Maluridae)
honeyeaters & Australian chats (Meliphagidae)
Thornbills and allies (Acanthizidae)
Superfamily Corvoidea
Australian robins (Petroicidae)
logrunners (Orthonychidae)
pseudo-babblers (Pomatostomidae)
diverse set of other birds:
crows & jays (Corvidae)
birds of paradise (Paradisaeidae)
wood-swallows, Australian magpies, butcherbirds, & currawongs (Artamidae)
magpie larks (Grallinidae)
bell magpies (Cracticidae)
whistlers, shrike-tit, thick heads, & pitohuis (Pachycephalidae)
This particular scenario is not universally accepted. However, it's apparent that many Australasian species evolved in Australia & nearby areas.
Bowerbirds - David Attenborough
Formation of New Guinean avian diversity across barriers and along elevational gradients by Knud Andreas Jonsson
Seminar - Poisonous birds and other stories of New Guinea fieldwork by Jack Dumbacher
| Songbirds
Escaped From Australasia, Conquered Rest of World - That
male cardinal singing his heart out in your backyard has ancestors that
left the neighborhood of Australia 45 million years ago. A
comprehensive
study of DNA from songbirds and their relatives by Barker et al. (2004)
shows that these birds, which account for almost half of all bird
species,
did not originate in Eurasia, as previously thought. Instead, their
ancestors
escaped from a relatively small area—Australasia (Australia, New
Zealand
and nearby islands) and New Guinea—about 45 million years ago and went
on to populate every other continent save Antarctica.
The movement of
the Australia/New
Guinea plate between 60 (a) and 20 (b) million years
ago
(Mya). When New Zealand
The order Passeriformes, or perching birds, includes all songbirds, such as robins, cardinals, blackbirds, house sparrows, house finches and crows. The group is further divided into birds that must learn their songs “true songbirds” (oscines) and those with the innate ability to sing the “correct” song. True songbirds account for 4,580 of the 6,000 known Passeriformes species. The true songbirds are currently divided into two groups: Passerida (3,477 species, among them many familiar backyard species) and Corvida (1,103 species, including crows and ravens). The two groups of true songbirds were thought to have separate origins, with Corvida originating in Australasia and the Passerida in Eurasia. The Passerida then supposedly spread from Eurasia to Africa, Australasia and the New World. But in examining the DNA sequences of two genes in all but one family (a closely related group, such as “crows and jays” or “warblers”) of passerine birds, Barker et al. (2004) made a startling discovery. “It was thought that the Passerida arose in Eurasia about 40 million years ago,” said Barker. “But we found that these birds fall into a group within the Corvida. That means all songbirds trace their origins to Australasia and New Guinea.” The Passerida differ from the Corvida because the Passerida somehow made it out of Australasia and New Guinea and onto the Asian mainland long before the Corvida, Barker said. Asia and Australasia are carried on separate plates in the Earth’s crust, and for many millions of years those plates were far apart. Around 45 million years ago, the ancestors of the Passerida dispersed to Asia—over more than 600 miles of open ocean--long before these two plates approached one another (as shown on map with locations of the continents 45 mybp). For some reason, however, ancestors of the Corvida didn’t make it until about 25 million years later, or 20 million years before today. At that time, Asia and Australia were much closer to each other, and island chains that could have allowed the Corvida ancestors to “island hop” to the mainland appeared, Barker said. “There are many endemic Corvida birds on the
Indonesian
island of Lombok but very few on Bali, the next island to the west,”
said
Barker. “And, sure enough, the line separating the Asian plate from the
Australasian plate runs between Bali and Lombok.
|
Neotropical
large number of endemics suggests an ancient & distinctive evolutionary history (Mayr 1964)
some endemic families have undergone extensive evolutionary radiation & contain many species, e.g.:
hummingbirds (Trochilidae) - 363 species (The secret life of hummingbirds)
ovenbirds (Furnariidae) - 242 species
antbirds, antshrikes, antwrens, antvireos, bushbirds, fire-eyes, & bare-eyes (Thamnophilidae) - 208 species
cotingas, bellbirds, becards, cocks-of-the-rock (Cotingidae) - 79 species
antthrushes (Formicariidae) & antpittas (Gralliidae)) - 56 species
manakins (Pipridae) - 52 species
woodcreepers (Dendrocolaptidae) - 52 species
puffbirds, nunlets, and nunbirds (Bucconidae) - 38 species
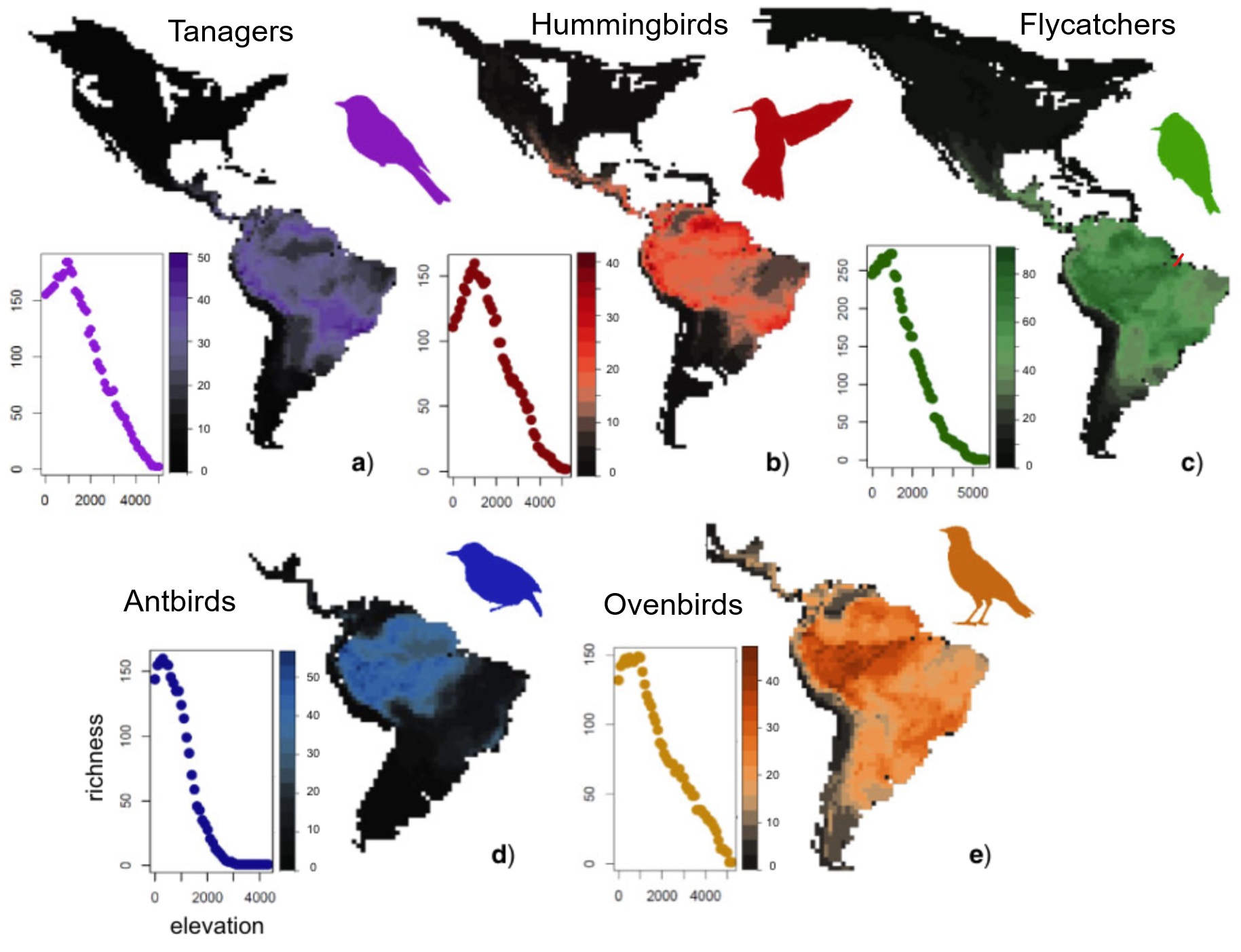
Five families of birds with high species richness in the Neotropics. Bird richness across regions and elevations in the
Neotropics. Richness for these five families declines toward temperate latitudes and at higher elevations: a) tanagers
(Thraupidae), b) hummingbirds (Trochilidae), c) tyrant flycatchers (Tyrannidae), d) antbirds (Thamnophilidae), and
e) ovenbirds (Furnariidae). Elevational panels indicate the global richness of the family at given elevation (in meters
above the sea level) (From: Machac 2020).

Black-fronted Nunbird
Photo © Arthur Grosset
tapaculos (Rhinocryptidae) - 30 species
jacamars (Galbulidae) - 17 species

Photograph courtesy Dr. Russell Barrow
other endemic families have undergone little radiation, e.g.:
potoos (Nyctibiidae) - 6 species
seedsnipes (Thinocoridae) - 4 species 
trumpeters (Psophiidae) - 3 species
plantcutters (Cotingidae) - 3 species
Hoatzin (Opisthocomidae) - 1 species (shown to the right)
Oilbird (Steatornithidae) - 1 species
in addition to these endemics, several groups are more recent arrivals from the Nearctic: vultures (Cathartidae), pigeons (Columbidae), jays (Corvidae), wrens (Troglodytidae), thrushes (Turdinae), blackbirds (Icterinae), & finches (Fringillidae)
several other groups have Pantropical distributions, including cuckoos (Cuculidae), parrots (Psittacidae), barbets (Capitonidae), & trogons (Trogonidae)
Neotropical forests are home to nearly 700 genera of birds.
Birds of Peru
Antarctic realm
The continent of Antarctica encompasses about 14.3 million km 2, with most covered by glacial ice. During the Antarctic summer, about 13,700 km 2 of rock and soil is exposed, particularly in the Antarctic Peninsula (Lee et al. 2017). Much of the ice-free area is along the edges of the continent, but other ice-free areas include mountain tops, cliffs, and valleys (Lee et al. 2017). These ice-free areas support breeding populations of 20 species of birds, including five species of penguins.
Species of birds that breed in Antarctica (Data from Harris et al. 2015).
Common name |
Scientific name |
Estimated global population (pairs) |
Emperor Penguin |
Aptenodytes forsteri |
238,000 |
Gentoo Penguin |
Pygoscelis papua |
387,000 |
Adélie Penguin |
Pygoscelis adeliae |
3,790,000 |
Chinstrap Penguin |
Pygoscelis antarctica |
2,650,000 |
Macaroni Penguin |
Eudyptes chrysolophus |
6,300,000 |
Wilson’s Storm-Petrel |
Oceanites oceanicus |
4 – 10,000,000 |
Black-belled Storm-Petrel |
Fregetta tropica |
160,000 |
Light-mantled Albatross |
Phoebetria palpebrata |
20,000 |
Southern Giant Petrel |
Macronectes giganteus |
50,000 |
Southern Fulmar |
Fulmarus glacialoides |
1,000,000 |
Antarctic Petrel |
Thalassoica antarctica |
3 - 7,000,000 |
Cape Petrel |
Daption capense |
670,000 |
Snow Petrel |
Pagodroma nivea |
1,300,000 |
Antarctic Prion |
Pachyptila desolata |
16,600,000 |
Imperial Shag |
Phalacrocorax atriceps |
13,000 |
Snowy Sheathbill |
Chionis albus |
10,000 |
Kelp Gull |
Larus dominicanus |
10 – 20,000 |
Antarctic Tern |
Sterna vittata |
36,700 |
South Polar Skua |
Catharacta maccormicki |
3000 - 7500 |
Brown Skua |
Catharacta antarctica |
3000 - 7500 |
Brown and South Polar skuas
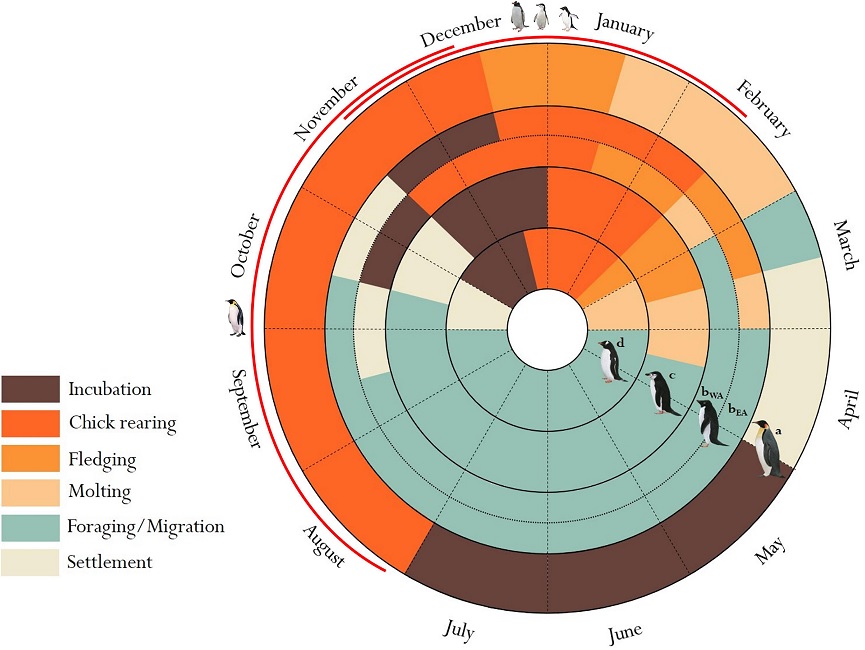
Life cycles of adults in four species of penguins that breed in Antarctica: (a) Emperor Penguins, (b) Adelie Penguins in East Antarctica (EA) and
West Antarctica (WA), (c) Chinstrap Penguins, and (d) Gentoo Penguins. Only Emperor Penguins incubate eggs during Antarctica's winter months (Figure from Handley et al. 2021).
Literature cited
Adler, G. H. 1994. Avifaunal diversity and endemism of tropical Indian Ocean islands. Journal of Biogeography 21: 85-95.
Avise, J.C. 2000. Phylogeography: the history and formation of species. Harvard University Press, Cambridge, MA.
Avise, J.C. and D. Walker. 1998. Pleistocene phylogeographic effects on avian populations and the speciation process. Proc. Roy. Soc. B 265:457-463.
Baker, A.J. 1991. A review of New Zealand ornithology. Current Ornithology 8:1-67.
Baker, A. J., S. L. Pereira, O. P. Haddrath, and K.-A. Edge. 2006. Multiple gene evidence for expansion of extant penguins out of Antarctica due to global cooling. Proceedings of the Royal Society B 273: 11-17.
Barker, F.K., A. Cibois, P. Schikler , J. Feinstein, and J. Cracraft. 2004. Phylogeny and diversification of the largest avian radiation. Proc. Natl. Acad. Sci. USA 101:11040-11045.
Beaman, M. 1994. Palearctic birds: a checklist of the birds of Europe, North Africa and Asia north of the foothills of the Himalayas. Harrier Publications, Stonyhurst. UK.
Bergstrom, D. M., and S. L. Chown. 1999. Life at the front: history, ecology and change on southern ocean islands. Trends in Ecology & Evolution 14: 472-477.
Blackburn, T. M., and R. P. Duncan. 2001. Establishment patterns of exotic birds are constrained by non-random patterns in introduction. Journal of Biogeography 28: 927-939.
Blackburn, T. M., J. L. Lockwood, and P. Cassey. 2009. Avian invasions: the ecology and evolution of exotic birds. Oxford University Press, Oxford, UK.
Blondel. J. and Cécile Mourer-Chauviré. 1998. Evolution and history of the western Palaearctic avifauna. Trends in Ecology and Evolution 13: 488-492.
Brooks, T., A. Balmford, N. Burgess, J. Fjeldsa, L. A. Hansen, J. Moore, C. Rahbek, and P. Williams. 2001. Toward a Blueprint for Conservation in Africa. BioScience 51: 613-624.
Bunce, M., M. Szulkin, H. R. L. Lerner, I. Barnes, B. Shapiro, A. Cooper, and R. N. Holdaway. 2005. Ancient DNA Provides New Insights into the Evolutionary History of New Zealand's Extinct Giant Eagle. PLoS Biology 3: e9.
Carnicer, J. and R. Diaz-Delgado. 2008. Geographic differences between functional groups in patterns of bird species richness in North America. Acta Oecologia 33: 253264.
Clarke, J. A., C. P. Tambussi, J. I. Noriega, G. M. Erickson, and R. A. Ketcham. 2005. Definitive fossil evidence for the extant avian radiation in the Cretaceous. Nature 433: 305-308.
Colwell et al. 2009. Peaks, plateaus, canyons, and craters: the complex geometry of simple mid-domain effect models. Evolutionary Ecology Research 11: 355370.
Cowie, R. H., and B. S. Holland. 2008. Molecular biogeography and diversification of the endemic terrestrial fauna of the Hawaiian Islands. Philosophical Transactions of the Royal Society B 363: 3363-3376.
Cracraft, J. 1974. Continental drift and vertebrate distribution. Ann. Rev. Ecol. Syst. 5:215-261.
Darlington, P.J. 1957. Zoogeography: the geographical distribution of animals. J. Wiley and Sons, New York, NY.
Ding, T.-S.
de Klerk et al. 2002. Patterns of species richness and narrow endemism of terrestrial bird species in the Afrotropical region. Journal of Zoology 256: 327-342.
Dorst, J. 1972. The evolution and affinities of the birds of Madagascar. In: Biogeography and ecology in Madagascar (R. Battistini and G. Richard-Vindard, eds.), pp. 615-627. Springer, Dordrecht, The Netherlands.
Dow, D. D. 1980. Communally breeding Australian birds with an analysis of distributional and environmental factors. Emu 80: 121-140.
Dumbacher, J. P., A. Wako, S. R. Derrickson, A. Samuelson, T. F. Spande, and J. W. Daly. 2004. Melyrid beetles (Choresine): A putative source for the batrachotoxin alkaloids found in poison-dart frogs and toxic passerine birds. Proc. Natl. Acad. Sci. 101: 15857-15860.
Dyer, E. E., P. Cassey, D. W. Redding, B. Collen, V. Franks, K. J. Gaston, K. E. Jones, S. Kark, C. D. L. Orme, and T. M. Blackburn. 2017. The global distribution and drivers of alien bird species richness. PLoS Biology 15: e2000942.
Ericson et al. 2003. Evolution, biogeography, and patterns of diversification in passerine birds. Journal of Avian Biology 34: 3-15.
Fabre, P. H., M. Irestedt, J. Fjeldså, R. Bristol, J. J. Groombridge, M. Irham, and K. A. Jønsson. 2012. Dynamic colonization exchanges between continents and islands drive diversification in paradise ‐flycatchers (Terpsiphone, Monarchidae). Journal of Biogeography 10: 1900-1918.
Feduccia, A. 1995. Explosive evolution in Tertiary birds and mammals. Science 267:637-638.
Feduccia, A. 2003. 'Big bang' for Tertiary birds? Trends in Ecology and Evolution 18:172-176.
Ficetola, G. F., F. Mazel, and W. Thuiller. 2017. Global determinants of zoogeographical boundaries. Nature Ecology & Evolution 1: 0089.
Filardi, C. E. and R. G. Moyle. 2005. Single origin of a pan-Pacific bird group and upstream colonization of Australia. Nature 438: 216-219.
Fitzpatrick, S. M., and W. F. Keegan. 2007. Human impacts and adaptations in the Caribbean Islands: an historical ecology approach. Earth and Environmental Science Transactions of the Royal Society of Edinburgh 98: 29-45.
Gill, F.B. 1995. Ornithology, second ed. W.H. Freeman and Co., New York, NY.
Gotelli, N. J., and L. G. Abele. 1982. Statistical distributions of West Indian land bird families. Journal of Biogeography 9: 421-435.
Handley, J., Rouyer, M.M., Pearmain, E.J., Warwick-Evans, V., Teschke, K., Hinke, J.T., Lynch, H., Emmerson, L., Southwell, C., Griffith, G. and Cárdenas, C.A., 2021. Marine important bird and biodiversity areas for penguins in Antarctica, targets for conservation action. Frontiers in Marine Science 7: 602972.
Harris, C.M., K. Lorenz, L. D. C. Fishpool, B. Lascelles, J. Cooper, N. R. Coria, J. P. Croxall, L. M. Emmerson, R. C. Fijn, W. L. Fraser, P. Jouventin, M. A. LaRue, Y. Le Maho, H. J. Lynch, R. Naveen, D. L. Fraser-Patterson, H.-U. Peter, S. Poncet, R. A. Phillips, C. J. Southwell, J. A. van Franeker, H. Weimerskirch, B. Wienecke, and E. J. Woehler. 2015. Important bird areas in Antarctica 2015. BirdLife International and Environmental Research & Assessment Ltd., Cambridge, UK.
Harshman, J., E. L. Braun, M. J. Braun, C. J. Huddleston, R. C. K. Bowie, J. L. Chojnowski, S. J. Hackett, K.-L. Han, R. T. Kimball, B. D. Marks, K. J. Miglia, W. S. Moore, S. Reddy, F. H. Sheldon, D. W. Steadman, S. J. Steppan, C. C. Witt, and T. Yuri. 2008. Phylogenomic evidence for multiple losses of flight in ratite birds. Proceedings of the National Academy of Sciences USA.
Hawkins, B. A., J. A. F. Diniz-Filho, C. A. Jaramillo, and S. A. Soeller. 2006. Post-Eocene climate change, niche conservatism, and the latitudinal diversity gradient of New World birds. Journal of Biogeography 33: 770-780.
Hawkins, B. A., J. A. F. Diniz-Filho, and S. A. Soeller. 2005. Water links the historical and contemporary components of the Australian bird diversity gradient.Journal of Biogeography 32: 1035–1042.
Hawkins, B. A., E. E. Porter, and J. A. F. Diniz-Filho. 2003. Productivity and history as predictors of the latitudinal diversity gradient of terrestrial birds. Ecology 84: 1608-1623.
Heinsohn, R. and M. C. Double. 2004. Cooperate or speciate: new theory for the distribution of passerine birds. Trends in Ecology and Evolution 19: 55-57.
Hurlbert, A. H. and J. P. Haskell. 2003. The effect of energy and seasonality on avian species richness and community composition. American Naturalist 161:83-97.
Hurlbert, A. H. and W. Jetz. 2007. Species richness, hotspots, and the scale dependence of range maps in ecology and conservation. Proceedings of the National Academy of Sciences 104: 13384-13389.
Irwin, M. T., P. C. Wright, C. Birkinshaw, B. L. Fisher, C. J. Gardner, J. Glos, S. M. Goodman, P. Loiselle, P., Rabeson, J. L. Raharison, and M. J. Raherilalao. 2010. Patterns of species change in anthropogenically disturbed forests of Madagascar. Biological Conservation 143: 2351-2362.
Jetz, W. and C. Rahbek. 2001. Geometric constraints explain much of the species richness pattern in African birds. Proceedings of the National Academy of Science 98:5661-5666.
Jønsson, K. A., P. H. Fabre, S. A. Fritz, R. S. Etienne, R. E. Ricklefs, T. B. Jørgensen, J. Fjeldså, C. Rahbek, P. G. Ericson, F. Woog, and E. Pasquet. 2012. Ecological and evolutionary determinants for the adaptive radiation of the Madagascan vangas. Proceedings of the National Academy of Sciences USA 109: 6620-6625.
Karr, J.R. 1990. Birds of tropical rainforest: comparative biogeography and ecology. Pp. 215-228 in Biogeography and ecology of forest bird communities (A. Keast, ed.). SPB Academic Publ., The Hague, Netherlands.
Kerr, K. C. R., M. Y. Stoeckle, C. J. Dove, L. A. Weigt, C. M. Francis, and P. D. N. Hebert. 2007. Comprehensive DNA barcode coverage of North American birds. Molecular Ecology Notes 7: 535543.
Klicka, J. and R.M. Zink. 1997. The importance of recent Ice Ages in speciation: a failed paradigm. Science 277:1666-1669.
Kosman et al. 2019. Conservation prioritization based on trait-based metrics illustrated with global parrot distributions. Diversity and Distributions 25: 1156-1165.
Lever, C. 1987. Naturalized birds of the world. Longman Scientific & Technical, Essex, England.
Levey, D.J. and F.G. Stiles. 1992. Evolutionary precursors of long distance migration: resource availability and movement patterns in Neotropical landbirds. American Naturalist 140:447-476.
Lijtmaer, D. A., D. C. R. Kerr, A. S. Barreira, P. D. N. Hebert, and P. L. Tubaro. 2011. DNA barcode libraries provide insight into continental patterns of avian diversification. PLoS ONE 6: e20744.
Lovei, G.L. 1989. Passerine migration between the Palearctic and Africa. Current Ornithology 6:143-174.
Lovette, I. J. and E. Bermingham. 1999. Explosive speciation in the New World Dendroica warblers. Proc. Roy. Soc. B 266: 1629-1636.
MacArthur, R.H., H. Recher, & M.L. Cody. 1966. On the relation between habitat selection and species diversity. Am. Nat. 100:319-332.
MacArthur, R.H. and E.O. Wilson. 1967. The theory of island biogeography. Princeton Univ. Press, Princeton, NJ.
Machac, A. 2020. The Dynamics of Bird Diversity in the New World. Systematic Biology 69: 11801199.
Martin-Albarracin, V. L., G. C. Amico, D. Simberloff, and M. A. Nuñez, 2915. Impact of non-native birds on native ecosystems: a global analysis. PLoS ONE 10: e0143070.
Mayr, E. 1946. History of the North American bird fauna. Wilson Bulletin 58:3-41.
Mayr, E. 1964. Inference concerning the Tertiary American bird faunas. Proc. Natl. Acad. Sci. 51:280-288.
McDowall, R.M., 2008. Process and pattern in the biogeography of New Zealand – a global microcosm? Journal of Biogeography 35: 197–212.
McGlone, M. S. 1989. The Polynesian settlement of New Zealand in relation to environmental and biotic changes. New Zealand Journal of Ecology 12: 115-129.
McKinney, M. and S. Kark. 2017. Factors shaping avian alien species richness in Australia vs Europe. Diversity and Distributions 23: 1334-1342.
Mengel, R.N. 1964. The probable history of species formation in some northern wood warblers (Parulidae). Living Bird 3:9-43.
Milá, B., J. E. McCormack, G. Castañeda, R. K. Wayne, and T. B. Smith. 2007. Recent postglacial range expansion drives the rapid diversification of a songbird lineage in the genus Junco. Proceedings of the Royal Society B 274:2645-2652.
Miyaka, C. Y., S. R. Matioli, T. Burke, and A. Wajntal. 1998. Parrot evolution and paleogeographical events: mitochondrial DNA evidence. Molecular Biology and Evolution 15: 544-551.
Mönkkönen, M. and P. Viro. 1997. Taxonomic diversity of the terrestrial bird and mammal fauna in temperate and boreal biomes of the northern hemisphere. J. Biogeogr. 24: 603–612.
Moreau, R.E. 1952. Africa since the Mesozoic: with particular reference to certain biological problems. Proc. Zool. Soc. London 121:869-913.
Morris, P., and F. Hawkins. 1998. Birds of Madagascar: a photographic guide. Yale University Press, New Haven, CT.
Moyle, R.G., R. T. Chesser, R. O. Prum, P. Schikler, and J. Cracraft. 2006. Phylogeny and evolutionary history of Old World suboscine birds (Aves: Eurylaimides). American Museum Novitates 3544: 1–22.
Neall, V. E., and S. A. Trewick. 2008. The age and origin of the Pacific islands: a geological overview. Philosophical Transactions of the Royal Society B 363: 3293-3308.
Newton, I. and L. Dale. 2001. A comparative analysis of the avifaunas of different zoogeographical regions. Journal of Zoology 254: 207-218.
O'Connor, J. 2022. Enantiornithes. Current Biology 32: R1166-R1172.
Oliveros et al. 2019. Earth history and the passerine superradiation. Proceedings of the National Academy of Sciences 116: 7916-7925.
Oong et al. 2026. Biogeographic Regions Within Indomalaya: An Integrated Approach Using Bird Distributions. Journal of Biogeography 53: e70155.
Porter, W. F.. 1994. Family Meleagrididae (Turkeys) in del Hoyo, J., Elliott, A., & Sargatal, J., eds. Handbook of the Birds of the World, Vol. 2. Lynx Edicions, Barcelona.
Prasetya, A. M. 2024. Historical Biogeography and Avian Speciation in the Indo-Australian Archipelago. Ph.D. dissertation, Australian National University, Canberra, Australia.
Price, J. P., and D. A. Clague. 2002. How old is the Hawaiian biota? Geology and phylogeny suggest recent divergence. Proceedings of the Royal Society B 269: 2429-2435.
Proctor, N.S. and P.J. Lynch. 1993. Manual of ornithology: avian structure and function. Yale Univ. Press, New Haven, CN.
Rabenold, K.N. 1993. Latitudinal gradients in avian species diversity and the role of long-distance migration. Current Ornithology 10:247-274.
Reddy, S., A. Driskell, D. L. Rabosky, S. J. Hackett, and T. S. Schulenberg. 2012. Diversification and the adaptive radiation of the vangas of Madagascar. Proceedings of the Royal Society B 279: 2062-2071.
Ritchie, J.C. 1982. Postglacial vegetation of Canada. Cambridge University Press, Oxford.
Roselaar, C. S., R. Sluys, M. Aliabadian, and P. G. M. Mekenkamp. 2007. Geographic patterns in the distribution of Palearctic songbirds. Journal of Ornithology 148: 271-280.
Samonds, K. E., L. R. Godfrey, J. R. Ali, S. M. Goodman, M. Vences, M. R. Sutherland, M. T. Irwin, and D. W. Krause. 2013. Imperfect isolation: factors and filters shaping Madagascar’s extant vertebrate fauna. PLoS ONE 8: e62086.
Sandel, B., L. Arge, B. Dalsgaard, R. G. Davies, K. G. Gaston, W. J. Sutherland, and J.-C. Svenning. 2011. The influence of Late Quaternary climate-change velocity on species endemism. Science334: 660-664.
Schulenberg, T. S. 2003. The radiations of passerine birds on Madagascar. In: The natural history of Madagascar (S. M. Goodman and J. P. Benstead, eds.), pp. 1130-1134. University of Chicago Press, Chicago, IL.
Selander, R.K. 1971. Systematics and speciation in birds. Pp. 57-147 in Avian Biology, vol. 1 (D.S. Farner and J.R. King, eds.). Academic Press, New York, NY.
Selvatti et al. 2022. Southern hemisphere tectonics in the Cenozoic shaped the pantropical distribution of parrots and passerines. Journal of Biogeography 49:17531766.
Shirihai, H. 2007. A complete guide to Antarctic wildlife: the birds and marine mammals of the Antarctic continent and the Southern Ocean. A&C Black Publishers, London, UK.
Sibley, C.G. and J.E. Ahlquist. 1985. The phylogeny and classification of the Australo-Papuan passerine birds. Emu 85:1-14.
Slack, K. E., C. M. Jones, T. Ando, G. L. Harrison, R. E. Fordyce, U. Arnason, and D. Penny. 2006. Early penguin fossils, plus mitochondrial genomes, calibrate avian evolution. Molecular Biology and Evolution 23:1144-1155.
Smith, N. A. 2016. Evolution of body mass in the Pan-Alcidae (Aves, Charadriiformes): the effects of combining neontological and paleontological data. Paleobiology 42: 8-26.
Smith, N. A. and J. A. Clarke. 2014. Systematics and evolution of the Pan-Alcidae (Aves, Charadriiformes). Journal of Avian Biology 45: 116.
Steadman, D. W. 1995. Prehistoric extinctions of Pacific island birds: biodiversity meets zooarchaeology. Science 267: 1123-1131.
Stiller et al. 2024. Complexity of avian evolution revealed by family-level genomes. Nature 629: 851860.
Tallis, J. 1990. Climate Change and Plant Communities. Academic Press, London.
Tennyson, A. J. D. 2010. The origin and history of New Zealand's terrestrial vertebrates. New Zealand Journal of Ecology 34: 6.
Toews, D. P. L. 2015. Evolution: a genomic guide to bird population history. Current Biology 25: R465-R467.
Valente, L., J. C. Illera, K. Havenstein, T. Pallien, R. S. Etienne, and R. Tiedemann. 2017. Equilibrium bird species diversity in Atlantic Islands. Current Biology 27: 1660-1666.
Valente et al. 2020. A simple dynamic model explains the diversity of island birds worldwide. Nature 579: 9296.
VanderWerf, E. A. 2012. Hawaiian Bird Conservation Action Plan. Pacific Rim Conservation, Honolulu, HI.
Walker, S., and A. Monks. 2017. New Zealand’s native land birds: status and change on the mainland from estimates of occupancy for 1969-1979 and 1999-2004. Parliamentary Commissioner for the Environment 2017, Wellington, New Zealand.
Wallace, A. R. 1876. The geographical distribution of animals. Macmillan, London.
Watson, D. M. 2005. Diagnosable versus distinct: evaluating species limits in birds. BioScience 55: 60-68.
Wiens, J.A. 1991. Distribution. Pp. 156-174 in The Cambridge Encylopedia of Ornithology (M. Brooke and T. Birkhead, eds.). Cambridge Univ. Press, New York, NY.
Welty, J.C. and L. Baptista. 1988. The life of birds, 4th ed. Saunders College Publishing, New York, NY.
Whittaker, R. J., J. M. Fernández-Palacios, T. J. Matthews, M. K. Borregaard, and K. A. Triantis. 2017. Island biogeography: taking the long view of nature’s laboratories. Science 357: eaam8326.
Widrig, K., and D. J. Field. 2022. The evolution and fossil record of Palaeognathous birds (Neornithes: Palaeognathae). Diversity 2022: 105.
Willson, M.F. 1976. The breeding distribution of North American migrant birds: a critique of MacArthur (1959). Wilson Bulletin 88:582-587.
Worthy, T. H., V. L. De Pietri, and R. P. Scofield. 2017. Recent advances in avian palaeobiology in New Zealand with implications for understanding New Zealand’s geological, climatic and evolutionary histories. New Zealand Journal of Zoology 44: 177-211.
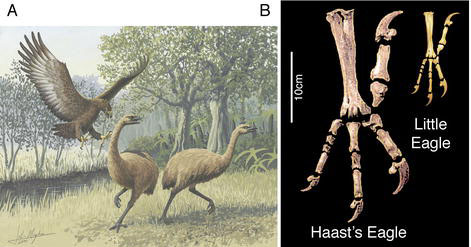

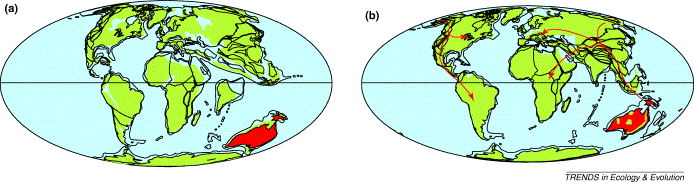
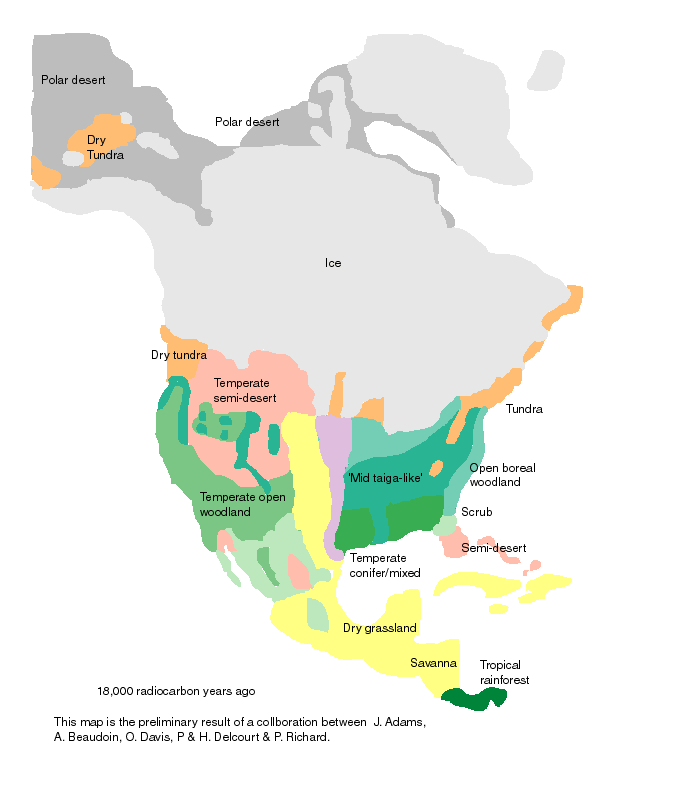{kind=link}