
Figure 1. Feathered (feather tracts) and unfeathered (apteria) areas of the avian integument (From: Chuong et al. 2000).

Figure 2. Cross-section through the skin of a bird or mammal (From: Lillywhite 2006).
BIO 554/754
Ornithology
Avian Integument
Epidermis | Integement modifications | Skin color | Skin color - structural | Claws | Rhamphotheca | Literature cited
The functions of bird skin are the same as for other vertebrates – to keep out pathogens and other potentially harmful substances, retain vital fluids and gases, and serve as a sensory organ. The continual renewal of the skin acts to repel parasitic microorganisms. The skin of birds also produces and supports feathers. With feathers, the skin also plays an important role in thermoregulation. Although largely covered by feathers, the integument is unfeathered on the beak, feet, and, in some species, other areas. In contrast to mammals, avian skin does not have sweat glands and sebaceous glands. Avian skin consists of two layers, the epidermis and dermis. The outer layer, the epidermis, is generally very thin and pliable. The dermis is thicker than the epidermis and contains blood vessels, fat deposits, nerves and free nerve endings, several types of neuroreceptors, and smooth muscles that move the feathers (Lucas and Stettenheim1972).
The epidermis, the most superficial layer of the skin, is thinner in birds than in mammals of comparable size, flexible, and smooth, and this is due, at least part, to selective pressures to minimize body weight for more efficient flight (Spearman 1966). The epidermis is thinnest in areas covered by feathers (both feather tracts and apteria; Figure 1) and thickest in exposed, featherless areas, including the covering of the beak (rhamphotheca) and feet (podotheca). The epidermis has two main layers – a superficial stratum corneum and a deeper strateus germinativium (Figure 2). The stratum corneum consists of flattened, keratinized cells. These cells are called keratinized because they contain a protein called keratin (and, specifically, beta keratin) that, along with extracellular lipids (fats) produced by epidermal cells, provide a tough, permeability barrier that prevents excessive evaporative water loss. The stratum corneum can be viewed as having a ‘brick-and-mortar’ organization, with the keratin-enriched cells forming the ‘bricks’ and the extracellular lipids the ‘mortar’ (Elias and Menon 1991). However, compared to reptiles and even mammals, cells in the stratum corneum have less keratin and, as a result, this barrier is less stringent and can facilitate evaporative cooling while retaining the capacity for facultative waterproofing (Menon et al. 1996). The high body temperatures of birds, increased heat production during flight, insulation by plumage and the lack of sweat glands, require a higher rate of evaporative cooling through a relatively "leaky" epidermal permeability barrier. Importantly, however, the relatively permeability of the avian epidermis can be modified. For example, Menon et al. (1996) found that, within 16 hours of water deprivation, adult Zebra Finches can reduce water loss via the epidermis by 50% by the rapid secretion of epidermal lipids. A similar ability to influence water loss by regulating secretion of epidermal lipids has been reported in larks (Haugen et al. 2003), House Sparrows (Passer domesticus; Muñoz-Garcia and Williams 2008), and the tropical Dusky Antbird (Cercomacra tyrannina; Muñoz-Garcia and Williams 2007).
Figure 1. Feathered (feather tracts) and unfeathered (apteria) areas of the avian integument (From: Chuong et al. 2000).
Figure 2. Cross-section through the skin of a bird or mammal (From: Lillywhite 2006).

Drawing showing the structure of avian skin
The epidermis consists of two layers, the upper stratum corneum keratinized or cornified flat cell sheets at the apical and the underlying stratum germinativum consisting of live cells. Nucleated cells of the stratum corneum form lamellae that are segregated by lipids. The dermis is thicker than the epidermis in many places of the body. Sensory nerve endings (Herbst corpuscles) occur throughout the dermis particularly associated with feather follicles. A thin elastic fiber layer separates the dermis from the subcutaneous layer that consists of randomly arranged collagen bundles and lipids. This adipose tissue is especially important for aquatic and migratory birds (From: Akat et al. 2022).
Besides forming a dynamic barrier that regulates water loss through the skin, epidermal lipids may also have antimicrobial properties and offer protection against ultraviolet light (Menon 1984). In addition, epidermal lipids are used for cosmetic coloration in the Japanese Crested Ibis (Nipponia nippon; Wingfield et al. 2000). Before breeding, the skin of the neck and head starts secreting a black substance that the ibises apply to their white plumage (Figure 3). The extent of the secretory skin area and how much of the plumage is covered by the ‘cosmetic’ varies among individuals and this variation plays a role in mate choice. Young birds do not produce the black secretion at all.

Figure 3. Plumages of the Japanese Crested Ibis. (a) Typical non-breeding plumage. (b) Daubing behavior; applying the pigment to plumage on the head, neck, and back. (c) ‘Cosmetically colored’ ibis. (d) After post-breeding molt (From: Wingfield et al. 2000).
Among many species of birds, the integument exhibits specialized modifications. For example, the skin on the head is unfeathered to varying degrees and distinctively colored in guineafowl, vultures, colies (Colius), and many storks, ibises, spoonbills, and cranes. The skin around the eyes is unfeathered and distinctively colored in other birds, such as cariamas, falcons, sheathbills (Chionis), parrots, cuckoos, broadbills, bare-eyes (Phlegopsis), lyrebirds (Menura), and helmet-shrikes (Prionops) (Stettenheim 2000). More generally, patches of bare skin, other than the bill and legs, can be found in birds belonging to at least 19 different orders and 62 families (Negro et al. 2006). Colored unfeathered areas on the head and necks of birds may be important in (1) intra- and intersexual communication, e.g., advertising status or quality, (2) thermoregulation, and (3) preventing the soiling of feathers for species that sometimes extend their heads into carcasses when feeding (e.g., vultures and condors).
Among birds with colorful skin (Figure 4), the coloration is due either to pigments, structural mechanisms in the epidermis, or to blood (and, specifically, hemoglogin in the red blood cells) in the superficial capillary network (Lucas 1970, Prum and Torres 2003).

Figure 4. Structurally colored ornaments of a sample of the piciform and passeriform birds examined: (A) Selenidera reinwardtii, (B) Ramphastos vitellinus, (C) Ramphastos toco, (D) Neodrepanis coruscans, (E) Philepitta castanea, (F) Myrmeciza ferruginea, (G) Gymnopithys leucapsis, (H) Procnias alba, (I) Perissocephalus tricolor, (J) Dyaphorophyia concreta, (K) Terpsiphone mutata and (L) Leucopsar rothschildi. A,F–J, reproduced with permission from VIREO; B,C,L, reproduced with permission from Kenneth Fink; H, reproduced with permission from Nate Rice; D, reproduced with permission from Steve Zack; K, reproduced with permission from Tom Schulenberg (From: Prum and Torres 2003).
Some species with bare skin on the head and neck alter skin color by changing blood flow to the area. For example, skin in the unfeathered areas of a Crested Caracaras (Polyborus plancus) head has a much denser supply of blood vessels than skin in feathered areas (Figures 5 and 6). Although they sometimes feed on carrion, the bare skin of caracaras is most important for thermoregulation. Caracaras are relatively large birds with generally dark plumage that are typically found in relatively hot areas. When their body temperature increases, blood flow to the areas of bare skin increases as vessels dilate. This increased blood flow causes the skin to become deeper red in color, but, most importantly, enhances the loss of heat across the bare skin. Bare skin on the head and neck likely serves a similar function for many species of birds because many such birds are relatively large, dark-plumaged birds that occur at low latitudes where heat dissipation may be of great importance (Negro et al. 2006).

Figure 5. A Crested Caracara showing unfeathered (a) and feathered (b) areas of the head (From: Negro et al. 2006).
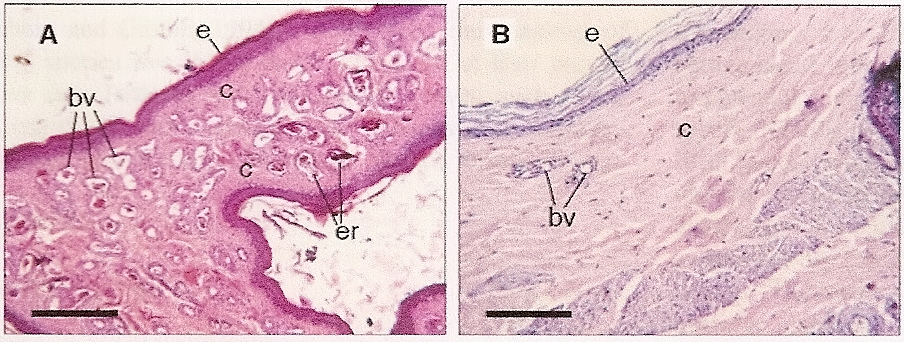
Figure 6. Micrographs of cross-sections of skin of a Crested Caracara, a species with unfeathered areas on the head. (A) Unfeathered area (bare skin) on the face, and (B) feather-covered area on the head. Note the greater number of blood vessels in the unfeathered skin. Scale bars: 25 µm. e, epidermis; c, collagen; er, erythrocytes; bv, blood vessels. (From: Negro et al. 2006).
For those species of birds with bare skin on the head and neck and where skin coloration is altered by changes in blood flow (‘flushing’), thermoregulation was almost certainly the primary selective factor. However, in some species, flushing occurs in contexts unrelated to thermoregulation, such as during courtship or agonistic encounters. For example, the skin of turkeys becomes redder when courting females and when engaged in agonistic encounters with other males. This suggests that ‘flushing’ can, for some species and in some contexts, also serve a signaling function. The ability to generate a deeper red coloration or maintain redder coloration for longer periods may be correlated with individual quality if doing so is energetically costly or potentially damaging to the body (Negro et al. 2006).
The colored skin of many birds is due to pigments, molecules that differentially absorb and emit wavelengths of visible light. Carotenoids are the pigments responsible for colorful skin (as well as feathers) in many birds, and typically generate a red, orange, or yellow hue. Birds cannot synthesize carotenoids so must acquire them in their diet. As a result, variation in carotenoid-based skin (or feather) coloration can provide conspecifics with information about individual quality and, specifically, the ability of different individuals to acquire a limited resource (Negro et al. 2002). For example, Red-legged Partridges (Alectoris rufa) have bills and eye rings (bare skin not feathers) that are reddish due to the presence of carotenoids and individuals with redder bills and eye rings are in better physical condition (Pérez-Rodríguez and Viñuela 2008). Similarly, the yellow-orange skin on the legs, feet, and ceres (skin at the base of the upper bill) of Eurasian Kestrels (Falco tinnunculus) is due to carotenoids and studies have revealed that male kestrels with more brightly colored skin are better hunters and have better quality territories (Casagrande et al. 2006).
For many birds, skin coloration is the result of optical interactions with biological nanostructures or, in other words, the microscopic structure of skin (Figure 7). Such structural colors occur in the skin, bill (ramphotheca), legs and feet (podotheca) in about 129 avian genera in 50 families from 16 avian orders. Structurally colored skin is present in more than 250 bird species, or about 2.5% of all bird species (Prum and Torres 2003). Examination of the color, anatomy, and nanostructure of structurally colored skin, ramphotheca, and podotheca from several different species of birds indicates that color, including ultraviolet, dark blue, light blue, green and yellow hues, is produced by coherent scattering (i.e., constructive interference) of light from arrays of parallel collagen fibers in the skin (dermis) (Figures 8 and 9). Scattering, in this case, simply means that light deviates from a straight path.
Visible light is, of course, composed of many colors of light with distinct wavelengths. Red light has a long wavelength (~700 nm), whereas violet and blue light has a much shorter wavelength (~400 nm). When visible light encounters particles with the same or larger diameter than its component wavelengths, those specific light photons are reflected. For example, particles that are 400 nm or slightly larger will selectively reflect blue light photons while allowing other light photons to pass. In the skin of birds light is reflected by collagen fibers (long, string-like protein molecules) that are arranged in a much more highly organized manner than in normal skin. For patches of skin that are a particular color (e.g., blue or green) all collagen fibers are the same thickness (Figure 8). As a result, each fiber scatters wavelengths of light that are in phase (Figure 10) and, therefore, are reinforced, producing very bright colors even more saturated than typical pigment-based colors. In addition to the reflection of light of certain wavelengths, skin structural coloration also requires a means to prevent the reflection or scattering of white light by deeper tissues below the color-producing nanostructures. In bird skin, this light-absorbing layer consists of a thick layer of melanin granules (melanosomes; Figure 7).

Figure 7. Light micrographs of structurally colored, white and pigmented bird skin showing differences between structurally colored and non-structurally colored skin in the thickness of collagen layers : (A) Chaemapetes unicolor, dark blue; (B) Numida meleagris, dark blue; (C) Tragopan temminckii, light blue; (D) Opisthocomus hoazin, dark blue; (E) Ramphastos vitellinus, dark blue; (F) Selenidera reinwardtii, green; (G) Procnias nudicollis, white leg skin; (H) Procnias nudicollis, structurally green throat skin and (I) Tragopan temminckii, red, lateral lappet patches. All specimens stained with Masson's trichrome, which stains collagen blue and cells red. All scale bars represent 100 µm, except in C, which represents 50 µm. Abbreviations: c, collagen macrofibrils; cc, collagenocytes; cp, capillaries; e, epidermis; m, melanosomes (From: Prum and Torres 2003).

Figure 8. Transmission electron micrographs showing the very regular nanostructured arrays of dermal collagen fibers from: (A) Oxyura jamaicensis, light blue; (B) Numida meleagris, dark blue; (C) Tragopan satyra, dark blue; (D) Tragopan caboti, dark blue; (E) Tragopan caboti, light blue; (F) Tragopan caboti, orange; (G) Syrigma sibilatrix, blue; (H) Ramphastos toco, dark blue; (I) Philepitta castanea, light blue; (J) Gymnopithys leucapsis, light blue; (K) Procnias nudicollis, green and (L) Terpsiphone mutata, dark blue. All images were taken at 30000 x. All scale bars represent 200 nm (From: Prum and Torres 2003).

Figure 9. Coherent scattering is differential interference or reinforcement of wavelengths scattered by multiple light-scattering objects (x, y). Coherent scattering of specific wavelengths is determined by the phase relationships among the scattered waves. Scattered wavelengths that are out of phase will cancel each other out, but scattered wavelengths that are in phase will be constructively reinforced and coherently scattered. Phase relationships of wavelengths scattered by two different objects (x and y) are given by the differences in the path lengths of light scattered by the first object (x: 1–1') and a second object (y: 2–2') as measured from planes perpendicular to the incident (a) and reflected (b) waves in the mean refractive index of the media (From: Prum and Torres 2003).
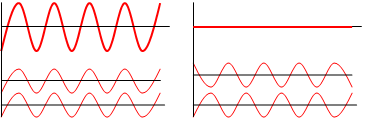
Figure 10. Left, In constructive interference (coherent scattering), two light waves (on bottom) are in phase so the combined waveform (top) is enhanced. Right, In destructive interference, two light waves (bottom) out of phase so they cancel out (top). (From: Wikipedia).
In some groups of birds (e.g., Phasianidae, Eurylaimidae, Cotingidae, Paradisaeidae, and Cnemophilidae), sexual selection is likely responsible for the structurally colored skin of males because most species are polygynous. In other groups of birds, including Ardeidae, Cariamidae, Bucerotidae, Ramphastidae, Meliphagidae, and Monarchidae, both sexes have integumentary structural colors, suggesting that such coloration may be important in both inter- and intrasexual communication (Prum and Torres 2003). Many birds with structurally colored skin are found in rainforests. For example, nearly all species with structurally colored skin in the orders Casuariiformes, Galliformes, Opisthocomiformes, Cuculiformes, Trogoniformes, Coraciiformes, Piciformes, and Passeriformes occur in tropical forests. The quality of ambient light in tropical forest habitats may favor the evolution of communication signals in the shorter wavelength portion of the visible spectrum (blue and green) and, if so, selection might favor structural colors because vertebrates, including birds, have no pigments that generate such colors (Prum and Torres 2003).

Surucua Trogon (Trogon surrucura)
Claws are found at the distal end of all toes of all birds and cover the bones of terminal phalanges. Some birds also have wing claws. The dorsal portion of claws is highly keratinized and calcified and is very hard. Claws are curved to varying degrees because the dorsal portion grows faster than the ventral portion. In addition to variation in curvature, claws also vary in relative length and pointedness (Stettenheim 2000). Among diving and swimming birds with webbed or partially webbed feet, such as gannets, waterfowl, and gulls, claws tend to be small, less curved, and flatter. At the extreme, grebes have very flat claws that increase foot surface area and aid in underwater propulsion. In contrast, birds that climb on vertical or nearly vertical surfaces, like woodpeckers and nuthatches, have sharply recurved and pointed claws to help grip the substrate (e.g., bark of a tree). Glen and Bennett (2007) examined claw morphology and placed terrestrial birds into six categories, with ground-dwelling birds having longer, less recurved claws and the amount of curvature increasing for birds that are increasingly arboreal in their foraging habits (Figure 11). Among raptorial birds that use their feet to capture and kill prey, claws (also called talons) are relatively long, highly recurved, and pointed.
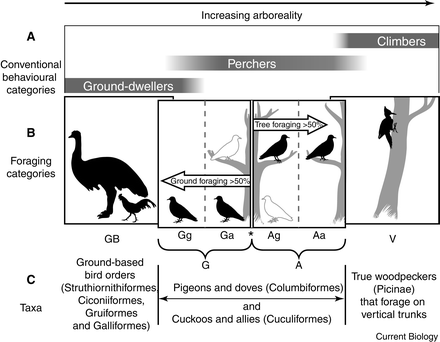
Figure 11. Categories (GB, Gg, Ga, Ag, Aa and V) of avian toe claws based the degree of ground or tree foraging; GB = ‘ground-based’ birds, limited to foraging on the ground; Gg = ‘dedicated ground foragers’; Ga = ‘predominantly ground foragers’; Ag = ‘predominantly arboreal foragers’; Aa = ‘dedicated arboreal foragers’; V = ‘vertical surface foragers’. Analysis of the toe claws of 249 species of birds revealed that claw curvature increases as tree foraging becomes more predominant (Glen and Bennett 2007).

Representative third pedal unguals showing typical claws for each of the three ecological groups. Flying taxa:
(a) Tauraco porphyreolophus (Purple-crested Tauraco) and (b) Psarocolius montezuma (Black Oropendola); Predatory
taxa: (c) Harpagus bidentatus (Double-toothed Kite) and (d) Aviceda leuphotes (Pacific Baza); Cursorial taxa:
(e) Dendragapus canadensis (Blue Grouse) and (f) Meleagris gallopavo (Wild Turkey). Predatory birds use their
highly recurved claws in prey capture while ground-dwelling birds typically have flat claws to ensure
that the claws do not interfere with locomotor ability. Claws of birds in the generalist flying category are more variable,
given much variation in their habitats and lifestyles (e.g., perching, climbing, arboreal species, species spending more time foraging on
the ground, etc.) (From: Hedrick et al. 2019).
Several species of birds representing 17 families of birds in eight orders Nightjars, and including herons, frigatebirds, and pratincoles, have pectinate middle claws with comb-like edges that are used for grooming and preening feathers (Figure 12; Stettenheim 2000, Moyer and Clayton 2003). For example, Fiery-necked Nightjars (Caprimulgus pectoralis) use their pectinated claw to groom their rather long rictal bristles (Jackson 2007), and nocturnally foraging nightjars, more generally, may use their pectinated claw to clean spider webs from their plumage (Masterson 1979).

Figure 12. A pectinate claw.
Wing claws can be found at the tip of the alular digit in several groups of birds, including loons, storks, owls, and some shorebirds, but are very small and non-functional. Young Hoatzins (Opisthocomus hoazin), however, have two, well-developed claws on each wing and use them for climbing shrubs and trees. These claws have an important function for Hoatzins, a Neotropical species, because they typically nest over water and nestlings sometimes jump from nests when threatened by a predator. Once in the water, they swim to nearby shrubs and trees and climbs upwards using their wing claws and feet. Young Hoatzins shed their wing claws when 70-100 days old (Thomas 1996).
Bird bills consist of bones that form the cores of the upper and lower mandibles. However, the outer surface and part of the inner surface of these bones are covered with a modified integument called the rhamphotheca (Figure 13). The epidermis of the rhamphotheca is relatively thick, hard, and consists of heavily cornified cells (Lucas and Stetteheim 1972). These cells produce beta-keratin like that found in avian scales and claws and calcium deposited between the keratin proteins generally make the rhamphotheca hard and strong (Homberger and Brush 1986, Bonser 1996). The epidermis is tightly bound to bone by a thin dermal layer that contains numerous collagen fibers. The dermis also contains sensory receptors, including Herbst and Grandry corpuscles that are sensitive to touch and vibration. The integument of the bill grows continually from the base and culmen, with growth directed rostrally so that there is continuous movement of the horny beak from base to tip. The rhamphotheca, particularly at the tomia, is worn away by abrasion from food and other materials and by friction where the upper and lower bills meet.
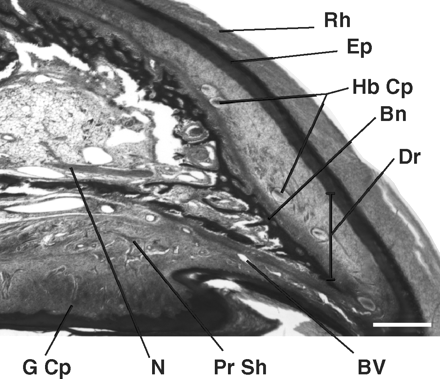
Figure 13. A sagittal section near midline of the upper beak of a 2-wk-old domestic chicken. The bill tip is to the right. The dorsal region shows the upper smooth portion of the bill covered by the rhamphotheca (Rh), whereas the ventral region shows the tomium or cutting edge of the upper bill. Beneath the rhamphotheca is the epidermis (Ep) that provides a constant supply of cellular material to form the outer, hardened covering of the beak. Internal to the epidermis is the dermis (Dr) layer that is the most heterogeneous of all tissue layers. It extends from the epidermal to bone layers (Bn). Prominent structures found in this region include mechanoreceptors (Herbst [Hb Cp]and Grandry [G Cp] corpuscles), blood vessels (BV), perineural sheaths, and free nerve endings, or nociceptors (pain) Scale bar, 400 µm. N = nerve; Pr Sh = perineural sheaths; (From: Kuenzel 2007).

Topographic anatomy of the rhamphotheca, showing rhamphothecal plates as well as nasolabial, culminolabial, and mentolabial grooves,
in (A) Northern Shoveler, (B) Common Loon, (C) Laysan Albatross, (D) Double-crested Cormorant, (E) Striated Heron, (F) Ring-billed Gull,
(G) Northern Flicker, and (H) American Crow. Where the loss of a rhamphothecal groove results in fusion of plates, those plates
are identified as composites (e.g., culminicorn + latericorn in Northern Shoveler) (From: Hieronymus and Witmer 2010).
The edges of the rhamphotheca (tomia) are relatively sharp in most species of birds. However, edges are finely serrated for straining small food particles in filter-feeding birds like flamingoes and some waterfowl. Species in a number of bird families, including Ardeidae, Cuculidae, Coraciidae, Picidae, and several others, have scopate tomia with very small (typically only 0.3-0.7 mm high and difficult to see without magnification) brush-like ridges that likely create friction and aid birds in capturing and holding food items (Figure 14; Gosner 1993). Tiny serrations along the tomia of one (just upper bill) or both tomia of some hummingbirds likely aid in capturing and holding insect prey and, in some species of nectar-robbers, piercing the base of flowers to gain access to nectar (Ornelas 1994; Figure 15). The tomia of fish-eating mergansers have numerous pointed projections that aid in capturing and holding their primary prey. Falcons and shrikes have single, sharp projections of the tomia on each side of their upper bill. Several authors have suggested that falcons use these ‘tomial teeth’ to break the neck of their prey, but there is little or no evidence to support this hypothesis. Rather, ‘tomial teeth’ are likely used to help firmly grip and pull flesh from prey (Csermely et al. 1998) and, for large falcons, to help grip and break long bones of wings and legs of smaller prey before swallowing (White et al. 2024).

Figure 14. Top, Scopate tomia of the fish-eating Ringed Kingfisher (Ceryle torquata). Bottom, scopate tomia of the insectivorous Laughing Kookaburra (Dacelo novaguineae). Tomia magnified 10X (From: Gosner 1993).
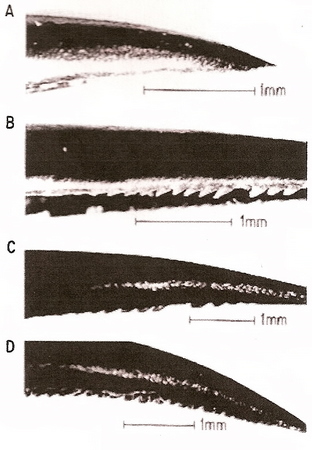
Figure 15. Upper mandibles of four species of hummingbirds showing a typical, smooth tomium (A, Violet Sabrewing, Campylopterus hemileucurus) plus the serrate tomia of the Sparkling Violetear (Colibri coruscans, B), Crowned Woodnymph (Thalurania ridgwayi, C), and Green-breasted Mango (Anthracothorax prevostii, D) (From: Ornelas 1994).
In some birds, including raptors (Falconiformes), owls (Strigiformes), parrots, cracids, and pigeons, the rhamphotheca at the base of the upper bill is called the cere (Figure 16; Stettenheim 1972). The cere is a thickened, often brightly colored portion of the integument that straddles the base of the nasal region. (Lucas 1979). Lucas and Stettenheim (1972) suggested that the cere may provide protection for the underlying elongated nasal fossa (cavities). However, because ceres are often brightly colored, individual variation in cere color may also convey information about individual quality. For example, the cere of male Montagu’s Harriers (Circus pygargus) reflects light at wavelengths corresponding to yellow-orange (500-600 nm), but also reflects ultraviolet (UV; 300-400 nm) wavelengths (Figure 17). Differences among males in the UV peak were found to be associated with differences in body mass and condition, suggesting UV reflectance conveys information about individual quality (Mougeot and Arroyo 2006).

Figure 16. The cere (C) of a Rock Pigeon located at the base of the upper bill. The arrow points to the external nares; (O), operculum (a small disc of cartilage centered in the nostril that keeps foreign objects out of the nasal cavity) (From: Purton 1988).
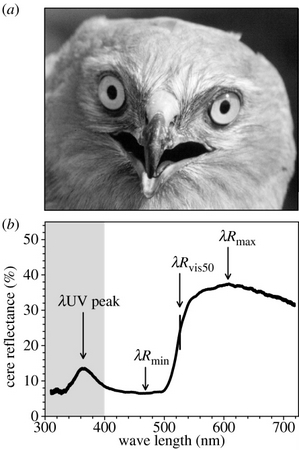
Figure 17. A male Montagu's Harrier (a) showing the cere (skin area above the beak and between the eyes); ( b) Reflectance pattern of a typical male cere showing peaks in the yellow-orange wavelengths (500-600 nm) and in the ultraviolet (300–400![]() nm) (From: Mougeot and Arroyo 2006).
nm) (From: Mougeot and Arroyo 2006).
Akat et al. 2022. Comparison of vertebrate skin structure at class level: A review. Anatomical Record 302: 3543-3608.
Bonser, B. H. C. 1996. Comparative mechanics of bill, claw and feather keratin in the Common Starling Sturnus vulgaris. Journal of Avian Biology 27: 175-177.
Casagrande , S., D. Csermely, E. Pini, V. Bertacche, and J. Tagliavini. 2006. Skin carotenoid concentration correlates with male hunting skill and territory quality in the kestrel Falco tinnunculus. Journal of Avian Biology 37: 190-196.
Chuong, C.-M., R. Chodankar, R. B. Widelitz, and T.-X. Jiang. 2000. Evo-Devo of feathers and scales: building complex epithelial appendages. Current Opinion in Genetics and Development 10: 449-456.
Csermely, D., L. Bertè, and R. Camoni. 1998. Prey killing by Eurasian Kestrels: the role of the foot and the significance of bill and talons. Journal of Avian Biology 29: 10-16.
Elias, P. M., and G. K. Menon. 1991. Structural and biochemical correlates of the epidermal permeability barrier. Advances in Lipid Research 24:1-26.
Glen, C. L., and M. B. Bennett. 2007. Foraging modes of Mesozoic birds and non-avian theropods. Current Biology 17: R911-R912.
Gosner, K. L. 1993. Scopate tomia: an adaptation for handling hard-shelled prey? Wilson Bulletin 105: 316-324.
Haugen, M., J. B. Williams, P. W. Wertz, and B. I. Tieleman. 2003. Lipids of the stratum corneum vary with cutaneous water loss among larks along a temperature-moisture gradient. Physiological and Biochemical Zoology 76: 907–917.
Hedrick et al 2019. Quantifying shape and ecology in avian pedal claws: The relationship between the bony core and keratinous sheath. Ecology and Evolution 9: 11545-11556.
Hieronymus, T. L. and L. M. Witmer. 2010. Homology and Evolution of Avian Compound Rhamphothecae. Ornithology 127: 590604.
Homberger, D. G., and A. H. Brush. 1986. Functional morphology and biochemical correlations of the keratinized structures of the African Gray Parrot, Psittacus erithacus (Aves). Zoomorphology 106: 103-114.
Jackson, H. D. 2007. Measurements and functions of the pectinated claws and rictal bristles of Fiery-necked Nightjars Caprimulgus pectoralis and some congeners. Ostrich 78: 641-643.
Kuenzel, W. J. 2007. Neurobiological basis of sensory perception: welfare implications of beak trimming. Poultry Science 86: 1273-1282.
Lillywhite, H. B. 2006. Water relations of tetrapod integument. Journal of Experimental Biology 209: 202-226.
Lucas, A. M. 1970. Avian functional anatomic problems. Fed. Proc. 29:1641-1648.
Lucas, A. M., and P. R. Stettenheim. 1972. Avian anatomy. Integument. Agriculture Handbook 362, U.S. Department of Agriculture, Washington, D.C.
Masterson, A. N. B. 1979. Of birds, the war and the future. Honeyguide 100: 7-11.
Menon, G. K. 1984. Glandular functions of avian integument: an overview. Journal of the Yamashina Institute for Ornithology 16:1–12.
Menon, G. K., P. F. A. Maderson, R. C. Drewes, L. F. Baptista, L. F. Price, and P. M. Elias. 1996. Ultrastructural organization of avian stratum corneum lipids as the basis for facultative cutaneous waterproofing. Journal of Morphology 227:1-13.
Mougeot, F., and B. E. Arroyo. 2006. Ultraviolet reflectance by the cere of raptors. Biology Letters 2: 173-176.
Moyer, B. R., and D. H. Clayton. 2003. Avian defenses against ectoparasites. In: Insect and bird interactions (H. F. van Emden and M. Rothschild, eds.), pp. 241-257. Intercept Ltd., Andover, UK.
Muñoz-Garcia, A., and J. B. Williams. 2007. Cutaneous water loss and lipids of the stratum corneum in Dusky Antbirds, a lowland tropical bird. Condor 109: 59-66.
Muñoz-Garcia, A., and J. B. Williams. 2008. Developmental plasticity of cutaneous water loss and lipid composition in stratum corneum of desert and mesic nestling House Sparrows. Proceedings of National Academy of Sciences USA 150:15611-15616.
Negro , J. J. , J. M. Grande , J. A Tella , J. Garrido , D. H ornero , J. A. Donázar , J. A. Sanchez-Zapata , J. R. Benítez , and M. Barcell . 2002 . An unusual source of essential carotenoids. Nature 416 : 807.
Negro, J. J., J. H. Sarasola, F. Fariñs, and I. Zorrilla. 2006. Function and occurrence of facial flushing in birds. Comparative Biochemistry and Physiology A 143: 78-84.
Ornelas, J. F. 1994. Serrate tomia: an adaptation for nectar robbing in hummingbirds? Auk 111: 703-710.
Pérez-Rodríguez, L., and J. Viñuela. 2008. Carotenoid-based bill and eye ring coloration as honest signals of condition: an experimental test in the Red-legged Partridge (Alectoris rufa). Naturwissenschaften 95: 821-830.
Prum, R. O., and R. Torres. 2003. Structural colouration of avian skin: convergent evolution of coherently scattering dermal collagen arrays. Journal of Experimental Biology 206: 2409-2429.
Purton, M. D. 1988. An ultrastructural study of the cere of the Domestic Pigeon (Columba livia). Journal of Anatomy 157: 43-56.
Spearman, R. I. C. 1966. The keratinization of epidermal scales, feathers, and hair. Biol. Rev. Cambridge Phil. Soc. 41: 59-96.
Stettenheim, P. 1972. The integument of birds. In: Avian biology, vol. II (D. S. Farner and J. R. King, eds.), pp. 1-63. Academic Press, New York, NY.
Stettenheim, P. R. 2000. The integumentary morphology of modern birds – an overview. American Zoologist 40: 461-477.
Thomas, B. T. 1996. Family Opisthocomidae (Hoatzin). In: Handbook of the birds of the world, vol. 3 (J. del Hoyo, A. Elliott, and J. Sargatal, eds.), pp. 23-32. Lynx Edicions, Barcelona, Spain.
White, C. M., N. J. Clum, T. J. Cade, and W. G. Hunt. 2024. Peregrine Falcon ( Falco peregrinus), version 1.1. In: Birds of the World (S. M. Billerman and M. G. Smith, Editors).. Cornell Lab of Ornithology, Ithaca, NY.
Wingfield, J. C., S. Ishii, M. Kikuchi, S. Wakabayashi, H. Sakai, N. Yamaguchi, M. Wada, and K. Chikatsuji. 2000. Biology of a critically endangered species, the Toki (Japanese Crested Ibis) Nipponia nippon. Ibis 142:1–11.